Lataa esitys
Esittely latautuu. Ole hyvä ja odota

1
Lääketieteen valmennuskurssi
BI5 BIOTEKNOLOGIA

3
1. Solun rakenne 1. Ensimmäiset solut olivat esitumaisia 2. Aitotumaiset solut ovat esitumaisia isompia ja monimutkaisempia 3. Tumallisissa soluissa on enemmän soluelimiä kuin esitumallisissa 4. Solut lähettävät ja vastaanottavat viestejä 5. Tehtävät

4
Ensimmäiset solut olivat esitumaisia
Prokaryootit Kehittyivät n. 3,8 mrd v.s. Pituudeltaan n mikrometriä Voivat muodostaa soluryhmiä Kuva kirjan sivulta 72

5
Aitotumaiset solut ovat esitumaisia isompia ja monimutkaisempia
Eukaryootit Alkueliöt, sienet, kasvit ja eläimet Kehittyivät esitumaisista n. 1,5 mrd v.s Endosymbioositeoria Mitokondriot ja viherhiukkaset kehittyivät yhteyttävistä bakteereista (muistuttavat kooltaan esitumaisia soluja) Pituudeltaan n. 0,1-0,01 mm Kuva kirjan sivulta 72
 Pituudeltaan n. 0,1-0,01 mm. Kuva kirjan sivulta 72.")
6
Nimeä rakenteet

7
Tumallisissa soluissa on enemmän soluelimiä kuin esitumallisissa
Kuva kirjan sivulta 72 ELÄINSOLU KASVISOLU

8
Solut lähettävät ja vastaanottavat viestejä
Aukkoliitos Siirtää solussa olevia ioneja ja molekyylejä naapurisoluun Kudoshormonit eli kasvutekijät Toimivat induktioaineina saavat solut mm. erilaistumaan Käynnistävät apoptoosin eli solukuoleman Kuva kirjan sivulta 72

9
Merkitse taulukkoon rasteilla, mitkä rakenteet löytyvät kustakin solutyypistä.
bakteerisolu eläinsolu kasvisolu sienisolu tuma dna tumakotelo solukalvo soluseinä kapseli solulima soluneste solunesterakkula

10
bakteerisolu eläinsolu kasvisolu sienisolu tuma x dna tumakotelo solukalvo soluseinä kapseli solulima soluneste (x) solunesterakkula
 solunesterakkula.")
11
2. Mikrobit 1. Mikrobit ovat mikroskooppisen pieniä 2. Aitotumaisiin mikrobeihin kuuluvat yksisoluiset alkueliöt sekä hiiva- ja homesienet 3. Arkit eivät ole bakteereita 4. Bakteereita on kaikkialla 5. Bakteerien viljely 6. Bakteerien lisääntyminen ja perimän muuttumistavat 7. Virusten rakenne 8. Prionit – oikea aminohappojärjestys – väärä proteiini 9. Tehtävät

12
Mikrobit ovat mikroskooppisen pieniä
Kuuluvat moneen eri eliökunnan ryhmään Myös virukset ovat mikrobeja Hyödylliset mikrobit (suolistobakteerit ja arkit) Haitalliset mikrobit Aiheuttavat sairauksia Kuva kirjan sivulta 72
 Haitalliset mikrobit. Aiheuttavat sairauksia. Kuva kirjan sivulta 72.")
13
Pöytävieraat (ei hyötyä eikä haittaa) Tuottajamikrobit
Syanobakteerit ja yhteyttävät yksisoluiset levät Kuluttajamikrobit (esim. tohvelieläin) Hajottajamikrobit Symbionttiset mikrobit Loismikrobit (esim. malarialoisio) Zoonoosi: mikrobin aiheuttama tauti, joka tarttuu eläimestä ihmiseen 14 A: Puutiainen (Ixodes ricinus) eli punkki on ulkoloinen, joka kiinnittyy isäntäänsä saadakseen verta. Puutiaiset elävät aluskasvillisuuden joukossa sekä heinikossa ja pensaikoissa. Tämä kumoaa vanhan kansan uskomuksen siitä, että punkit pudottautuvat leppäpuista. Puutiaiset toimivat sekä puutiaisaivokuumeen että borrelioosin levittäjinä. Pohjois-Karjalassa on tavattu siperianpunkkia (Ixodes persulcatus), joka levittää tappavaa aivokalvontulehdusta. Puutiaisen elämänkierto toukasta aikuiseksi kestää noin 2–3 vuotta, ja sen läpikäyminen vaatii kolme veriateriaa. Jos tätä ei joinakin vuosina ole tarjolla, punkin elinikä voi venähtää jopa 16 vuodeksi. Punkki ei varsinaisesti ime verta, vaan oikeammin juo ihohaavaan tihkuvan veren. Veriateria kestää useita päiviä ja tänä aikana punkki ”palauttaa” osan syödystä materiaalista takaisin isäntäeläimen vereen siirtäen näin myös mahdollisia taudinaiheuttajia, kuten borreliabakteerin. Tunnetuin puutiaisen levittämä bakteeri on Borrelia burgdorferi. Tämä bakteeri elää sulassa sovussa pikkujyrsijöiden elimistössä. Puutiaiseen borreliabakteeri siirtyy puutiaisen imiessä verta mm. hiirestä tai myyrästä. Kissa on myös varsin vastustuskykyinen borreliabakteerille, mutta koirille ja ihmiselle voi kehittyä vakava borrelioosisairaus 14 B: Malarian aiheuttaaPlasmodium-sukuun kuuluva alkueläin, joka siirtyy malariatartunnan saaneesta ihmisestä toiseen Anopheles-lajin hyttysen piston välityksellä. Alkueläin kulkeutuu hyttysen piston jälkeen maksaan ja sieltä edelleen veren punasoluihin. Neljä plasmodilajia aiheuttaa ihmisen malarian:Plasmodium falciparum, P. vivax, P. ovale ja P. malariae. Apinan P. Knowlesi voi aiheuttaa malarian myös ihmiselle, sitä on raportoitu Malesiasta. Malaria voi tarttua joskus, tosin harvoin, saastuneiden neulojen tai verensiirron välityksellä, kuten muutkin veren välityksellä tarttuvat infektiot. Sikiö voi saada malarian äidistään istukan kautta. Malarian tyypillisimmät oireet ovat toistuvat, vilunväreillä (horkka) alkavat kuumekohtaukset ja runsas hikoilu kuumeen laskiessa. Malariakuume voi usein, erityisesti alkuvaiheessa, olla jatkuvaa.
 Hajottajamikrobit. Symbionttiset mikrobit. Loismikrobit (esim. malarialoisio) Zoonoosi: mikrobin aiheuttama tauti, joka tarttuu eläimestä ihmiseen. 14 A: Puutiainen (Ixodes ricinus) eli punkki on ulkoloinen, joka kiinnittyy. isäntäänsä saadakseen verta. Puutiaiset elävät aluskasvillisuuden joukossa. sekä heinikossa ja pensaikoissa. Tämä kumoaa vanhan kansan uskomuksen. siitä, että punkit pudottautuvat leppäpuista. Puutiaiset toimivat sekä puutiaisaivokuumeen. että borrelioosin levittäjinä. Pohjois-Karjalassa on tavattu. siperianpunkkia (Ixodes persulcatus), joka levittää tappavaa aivokalvontulehdusta. Puutiaisen elämänkierto toukasta aikuiseksi kestää noin 2–3 vuotta, ja sen läpikäyminen vaatii kolme veriateriaa. Jos tätä ei joinakin vuosina ole. tarjolla, punkin elinikä voi venähtää jopa 16 vuodeksi. Punkki ei varsinaisesti. ime verta, vaan oikeammin juo ihohaavaan tihkuvan veren. Veriateria kestää. useita päiviä ja tänä aikana punkki palauttaa osan syödystä materiaalista. takaisin isäntäeläimen vereen siirtäen näin myös mahdollisia taudinaiheuttajia, kuten borreliabakteerin. Tunnetuin puutiaisen levittämä bakteeri on. Borrelia burgdorferi. Tämä bakteeri elää sulassa sovussa pikkujyrsijöiden elimistössä. Puutiaiseen borreliabakteeri siirtyy puutiaisen imiessä verta mm. hiirestä tai myyrästä. Kissa on myös varsin vastustuskykyinen borreliabakteerille, mutta koirille ja ihmiselle voi kehittyä vakava borrelioosisairaus. 14 B: Malarian aiheuttaaPlasmodium-sukuun kuuluva alkueläin, joka siirtyy. malariatartunnan saaneesta ihmisestä toiseen Anopheles-lajin hyttysen piston. välityksellä. Alkueläin kulkeutuu hyttysen piston jälkeen maksaan ja sieltä edelleen veren punasoluihin. Neljä plasmodilajia aiheuttaa ihmisen. malarian:Plasmodium falciparum, P. vivax, P. ovale ja P. malariae. Apinan P. Knowlesi voi aiheuttaa malarian myös ihmiselle, sitä on raportoitu Malesiasta. Malaria voi tarttua joskus, tosin harvoin, saastuneiden neulojen tai verensiirron. välityksellä, kuten muutkin veren välityksellä tarttuvat infektiot. Sikiö voi saada malarian äidistään istukan kautta. Malarian tyypillisimmät. oireet ovat toistuvat, vilunväreillä (horkka) alkavat kuumekohtaukset ja runsas. hikoilu kuumeen laskiessa. Malariakuume voi usein, erityisesti alkuvaiheessa, olla jatkuvaa.")
14
Aitotumaisiin mikrobeihin kuuluvat yksisoluiset alkueliöt sekä hiiva- ja homesienet
Yksinkertainen tumallinen solurakenne, erilaistuneiden kudosten puute Yksisoluiset levät Alkueläimet: toisenvaraisia ja liikkuvia Hiivat Yksisoluiset sienet Energia joko aerobisesti tai anaerobisesti Homesienet Rihmamainen kasvu (hyyfi eli rihma) Yksi tai useampi solu (soluseinä kitiiniä) Sopivan kostea ja hapekas elinympäristö Myrkkyjen eritys (jotkin lajit) 15 B: Alexander Fleming (1881–1955) on penisilliinin keksijä. Flemingillä oli laboratoriossaan bakteeriviljelymaljoja, jotka hän oli jättänyt lomansa ajaksi laboratorioon. Palattuaan hän huomasi erääseen bakteeriviljelymaljaan vahingossa päässeen homepesäkkeen ja sen ympärillä kehän, jossa maljassa viljellyt stafylokokit eivät kasvaneetkaan. Hän alkoi tutkia maljaa ja totesi, että Penicillium-sukuun kuu.luva home eritti kasvupaikkansa ympärille ainetta, joka aiheutta stafylokokeissa sekä kasvun estymistä että bakteerien hajoamista. Aineelle annettiin nimi penisilliini sitä valmistavan homeen mukaan. Howard Florey ja Ernst Boris Chain eristivät ja kiteyttivät penisilliinin 10 vuotta myöhemmin, jonka jälkeen se otettiin kliiniseen käyttöön. Pensselihomeita tavataan yli 500 lajia ja niitä on vaikeaa tunnistaa ja erottaa toisistaan. Penicillium on yleisin sisätiloissa todettava home. Sitä todetaan pieninä pitoisuuksina lähes kaikissa rakennuksissa (80 %:ssa pölynäytteistä ja 47 %:ssa ilmanäytteistä). Kosteusvaurion yhteydessä se voi alkaa kasvaa rakenteissa ja tuolloin se voi myös aiheuttaa merkittäviä terveyshaittoja. Penicilliumia pidetään ensilinjan homeena eli tuoreeseen kosteusvaurioon viittaavana mikrobina. Pensselihomeita esiintyy myös elintarvikkeissa, kuten leivässä.
 Yksi tai useampi solu (soluseinä kitiiniä) Sopivan kostea ja hapekas elinympäristö. Myrkkyjen eritys (jotkin lajit) 15 B: Alexander Fleming (1881–1955) on penisilliinin keksijä. Flemingillä oli. laboratoriossaan bakteeriviljelymaljoja, jotka hän oli jättänyt lomansa ajaksi. laboratorioon. Palattuaan hän huomasi erääseen bakteeriviljelymaljaan vahingossa. päässeen homepesäkkeen ja sen ympärillä kehän, jossa maljassa viljellyt. stafylokokit eivät kasvaneetkaan. Hän alkoi tutkia maljaa ja totesi, että. Penicillium-sukuun kuu.luva home eritti kasvupaikkansa ympärille ainetta, joka aiheutta stafylokokeissa sekä kasvun estymistä että bakteerien hajoamista. Aineelle annettiin nimi penisilliini sitä valmistavan homeen mukaan. Howard Florey ja Ernst Boris Chain eristivät ja kiteyttivät penisilliinin 10. vuotta myöhemmin, jonka jälkeen se otettiin kliiniseen käyttöön. Pensselihomeita tavataan yli 500 lajia ja niitä on vaikeaa tunnistaa ja erottaa. toisistaan. Penicillium on yleisin sisätiloissa todettava home. Sitä todetaan. pieninä pitoisuuksina lähes kaikissa rakennuksissa (80 %:ssa pölynäytteistä. ja 47 %:ssa ilmanäytteistä). Kosteusvaurion yhteydessä se voi alkaa kasvaa. rakenteissa ja tuolloin se voi myös aiheuttaa merkittäviä terveyshaittoja. Penicilliumia. pidetään ensilinjan homeena eli tuoreeseen kosteusvaurioon viittaavana. mikrobina. Pensselihomeita esiintyy myös elintarvikkeissa, kuten. leivässä.")
15
Arkit eivät ole bakteereita
Solukalvon kemiallinen rakenne poikkeaa bakterien ja aitotumaisten soluista (soluseinä proteiinia) Geeneissä introneita Haastavat elinympäristöt (esim. kuumat lähteet, suolajärvet) Arkit ja muut lajit keskenään mutualistisia Kuva kirjan sivulta 72
 Geeneissä introneita. Haastavat elinympäristöt (esim. kuumat lähteet, suolajärvet) Arkit ja muut lajit keskenään mutualistisia. Kuva kirjan sivulta 72.")
16
Bakteereita on kaikkialla
Omavaraiset eli autotrofiset bakteerit Fotosynteettiset bakteerit (esim. syanobakteerit) Rihmamaiset muodostelmat Yhteyttävät Auringon valon avulla Kemosynteettiset (kemiautotrofiset) bakteerit (esim. maaperän nitrifioivat bakteerit) Energia yhteyttämiseen ammoniakki-, rauta- tai rikkiyhdisteitä hapettamalla Toisenvaraiset bakteerit Orgaaniset yhdisteet muilta eliöiltä Kuva kirjan sivulta 72
 Rihmamaiset muodostelmat. Yhteyttävät Auringon valon avulla. Kemosynteettiset (kemiautotrofiset) bakteerit (esim. maaperän nitrifioivat bakteerit) Energia yhteyttämiseen ammoniakki-, rauta- tai rikkiyhdisteitä hapettamalla. Toisenvaraiset bakteerit. Orgaaniset yhdisteet muilta eliöiltä. Kuva kirjan sivulta 72.")
17
Grampositiivisten bakteerien erityispiirteitä:
– paksu, violetiksi värjäytyvä soluseinä – soluseinän rakenne yksinkertainen – soluseinä koostuu peptidoglykaaneista, teikoiinihapoista ja lipoteikoiinihapoista, jotka vaikuttavat mm. ionitasapainon säätelyyn – polysakkarideja – herkkiä antibiooteille Gramnegatiivisen bakteerien erityispiirteitä: – ohut, punaiseksi värjäytyvä soluseinä (jälkivärjäyksellä) – soluseinän rakenne monimutkainen – eivät ole herkkiä antibiooteille – soluseinässä lipoproteiineja ja lipopolysakkarideja sisältäväkerros peptidoglykaanin päällä, mikä suojaa mm. haitallisilta aineilta
 – soluseinän rakenne monimutkainen. – eivät ole herkkiä antibiooteille. – soluseinässä lipoproteiineja ja lipopolysakkarideja sisältäväkerros peptidoglykaanin. päällä, mikä suojaa mm. haitallisilta aineilta.")
19
A = basilli B = diplokokki C = streptokokki D = stafylokokki
E = spirilli 16 B: Stafylokokit eli rypälebakteerit (grampositiiviset bakteerit). Staphylococcus aureus -bakteeri kasvaa yleisesti ihmisen ja monien eläinten iholla ja limakalvoilla. Osa sen bakteerikannoista muodostaa kasvaessaan myrkkyä, joka aiheuttaa ruokamyrkytystä. Tämä toksiini on lämpöä kestävä eikä se tuhoudu tavallisessa kypsennyksessä. Siksi on tärkeää, että huolehtii puhtaudesta ja käsien ihon kunnosta sekä että työskentelee puhtaasti. Useille antibiooteille resistenttejä kantoja nimitetään MRSA-bakteereiksi (metisilliiniresistentti Staphylococcus aureus). Vakavia stafylokokkiinfektioita on perinteisesti hoidettu penisilliinin sukuisilla antibiooteilla. Jotkut stafylokokit ovat kuitenkin kehittyneet vastustuskykyiseksi näille tavallisille stafylokokkiantibiooteille. Streptokokit eli ketjukokit (grampositiiviset bakteerit) kuuluvat ihmisen suun ja nielun normaalikasvustoon. Monet niistä ovat taudinaiheuttajia. Streptokokkibakteerin tai virusten aiheuttama nielutulehdus on vaikea erottaa toisistaan. Streptokokki ja virukset aiheuttavat hyvin samankaltaisen taudinkuvan. Nielunäytteellä saadaan usein selvyys, onko tauti streptokokin vai viruksen aiheuttama.
. Staphylococcus. aureus -bakteeri kasvaa yleisesti ihmisen ja monien eläinten iholla ja. limakalvoilla. Osa sen bakteerikannoista muodostaa kasvaessaan myrkkyä, joka aiheuttaa ruokamyrkytystä. Tämä toksiini on lämpöä kestävä eikä se tuhoudu. tavallisessa kypsennyksessä. Siksi on tärkeää, että huolehtii puhtaudesta. ja käsien ihon kunnosta sekä että työskentelee puhtaasti. Useille antibiooteille. resistenttejä kantoja nimitetään MRSA-bakteereiksi (metisilliiniresistentti. Staphylococcus aureus). Vakavia stafylokokkiinfektioita on perinteisesti. hoidettu penisilliinin sukuisilla antibiooteilla. Jotkut stafylokokit ovat. kuitenkin kehittyneet vastustuskykyiseksi näille tavallisille stafylokokkiantibiooteille. Streptokokit eli ketjukokit (grampositiiviset bakteerit) kuuluvat ihmisen. suun ja nielun normaalikasvustoon. Monet niistä ovat taudinaiheuttajia. Streptokokkibakteerin tai virusten aiheuttama nielutulehdus on vaikea erottaa. toisistaan. Streptokokki ja virukset aiheuttavat hyvin samankaltaisen. taudinkuvan. Nielunäytteellä saadaan usein selvyys, onko tauti streptokokin. vai viruksen aiheuttama.")
20
O = bakteerien ryhmissä tavattava itiötyyppi
F = siima G = karva H = kapseli I = soluseinä J = solukalvo K = solulima L = ribosomit M = kromosomi N = plasmidi O = bakteerien ryhmissä tavattava itiötyyppi Bakteerien liikkuminen: Useilla bakteereilla on kyky liikkua kohti hyödyllistä suuntaa, kuten kohti ravinteikkaampaa ympäristöä tai valoa sekä myös poispäin itselleen vaarallisesta suunnasta. Vaihtelevassa ympäristössä solu ui kauemmin suotuisaan kuin epäsuotuisaan suuntaan, jolloin kokonaisliike tapahtuu kohti suotuisampaa suuntaa. Homogeenisessä ympäristössä solu ui sekunnin kerrallaan yhteen suuntaan ja vaihtaa sen jälkeen suuntaa, jolloin bakteerin liike on sattumanvaraista. Useilla bakteereilla on liikkumista varten siimat, mutta osa liikkuu myös liman avulla.

21
Bakteerien viljely Sopivat kasvatusolosuhteet
Petrimaljalla oleva agar-hyytelö Kuva kirjan sivulta 72

22
Bakteerien lisääntyminen ja perimän muuttumistavat
Suvuton jakautuminen Epäsuotuisissa olosuhteissa bakteerit lepoitiöitä Bakteerin perimässä useita plasmideja, mutta vain yksi kromosomi Bakteerien perimän muutokset Mutaatiot Transformaatio Dna:ta bakteerisoluun ulkopuolelta Transduktio Bakteriofagi-viruksen perimän mukana aiempien isäntäsolujen perimää bakteerisoluun Konjugaatio Bakteeri yhtyy viereiseen bakteeriin piluksen avulla Kuva kirjan sivulta 72

23
Bakteerien lisääntyminen

25
Virusten rakenne Ei omaa solurakennetta eikä aineenvaihduntaa
Tarvitsee isäntäsolun lisääntyäkseen (loisia) Eri eliöillä on omat viruksensa Perimä dna:ta tai rna:ta Myös nukleiinihapoista ja proteiineista koostuvat virukset katsotaan yleensä kuuluviksi mikrobeihin, vaikka viruksilla ei olekaan omaa solurakennetta ja aineenvaihduntaa, vaan ne tarvitsevat soluja elinkiertoonsa. Bakteerisolujen viruksia sanotaan faageiksi tai bakteriofaageiksi. Ovatko virukset ja faagit loisia vai itsekkäiden geenien pakkauksia? Faageja voi pitää samanlaisina loisina kuten aitotumaistenkin solujen viruksia, mutta faagit tarvitsevat nimenomaan bakteerisoluja elinkiertoonsa. Faagit ja virukset kuitenkin eroavat eliöiden 'tavanomaisista' loisista, kuten ihmisen lapamadosta siinä, että ne eivät välttämättä säästä isäntäsoluaan. Sen sijaan esimerkiksi lapamadon, kannattaa olla tuhoamatta isäntäänsä ainakin siinä vaiheessa, jolloin se on isännästään riippuvainen, sillä muutenhan loinenkin tuhoutuu. Sekä faagien että virusten olemassaolo on nähtävissä myös geenin itsekkyyden näkökulmasta; kaikki turhat elintoiminnat on karsittu mahdollisimman vähiin, jotta geenit voivat tuottaa ja levittää itsestään kopioita mahdollisimman tehokkaasti isäntäsolun kustannuksella. 22 A: Adenovirukset ovat saaneet nimensä kitarisojen mukaan, joista virus on ensimmäisenä eristetty. Adenovirukset aiheuttavat hengitystieinfektioita, ripulia ja silmätulehduksia. Influenssavirukset ovat yleisiä hengitystieinfektioita aiheuttavia viruksia. Ne jaetaan A-, B- ja C-tyyppeihin. Näistä A-tyyppi on yleisin taudinaiheuttaja. Influenssavirus A on mitä todennäköisimmin lähtöisin linnuista. Linnut pystyvät kantamaan tätä virusta ilman minkäänlaisia näkyviä oireita. Aasiassa, mistä useimmat influenssapandemiat ovat lähtöisin, linnut ja siat ovat läheisessä yhteydessä, koska esimerkiksi ankkoja ja sikoja pidetään samoilla alueilla kotieläiminä. Sioissa viruksesta kehittyy muoto, joka voi tarttua myös ihmiseen, minkä jälkeen influenssavirus voi tarttua myös ihmisestä toiseen.
 Eri eliöillä on omat viruksensa. Perimä dna:ta tai rna:ta. Myös nukleiinihapoista ja proteiineista koostuvat virukset katsotaan yleensä kuuluviksi mikrobeihin, vaikka viruksilla ei olekaan omaa solurakennetta ja aineenvaihduntaa, vaan ne tarvitsevat soluja elinkiertoonsa. Bakteerisolujen viruksia sanotaan faageiksi tai bakteriofaageiksi. Ovatko virukset ja faagit loisia vai itsekkäiden geenien pakkauksia Faageja voi pitää samanlaisina loisina kuten aitotumaistenkin solujen viruksia, mutta faagit tarvitsevat nimenomaan bakteerisoluja elinkiertoonsa. Faagit ja virukset kuitenkin eroavat eliöiden tavanomaisista loisista, kuten ihmisen lapamadosta siinä, että ne eivät välttämättä säästä isäntäsoluaan. Sen sijaan esimerkiksi lapamadon, kannattaa olla tuhoamatta isäntäänsä ainakin siinä vaiheessa, jolloin se on isännästään riippuvainen, sillä muutenhan loinenkin tuhoutuu. Sekä faagien että virusten olemassaolo on nähtävissä myös geenin itsekkyyden näkökulmasta; kaikki turhat elintoiminnat on karsittu mahdollisimman vähiin, jotta geenit voivat tuottaa ja levittää itsestään kopioita mahdollisimman tehokkaasti isäntäsolun kustannuksella. 22 A: Adenovirukset ovat saaneet nimensä kitarisojen mukaan, joista virus. on ensimmäisenä eristetty. Adenovirukset aiheuttavat hengitystieinfektioita, ripulia ja silmätulehduksia. Influenssavirukset ovat yleisiä hengitystieinfektioita aiheuttavia viruksia. Ne jaetaan A-, B- ja C-tyyppeihin. Näistä A-tyyppi on yleisin taudinaiheuttaja. Influenssavirus A on mitä todennäköisimmin lähtöisin linnuista. Linnut. pystyvät kantamaan tätä virusta ilman minkäänlaisia näkyviä oireita. Aasiassa, mistä useimmat influenssapandemiat ovat lähtöisin, linnut ja siat ovat. läheisessä yhteydessä, koska esimerkiksi ankkoja ja sikoja pidetään samoilla. alueilla kotieläiminä. Sioissa viruksesta kehittyy muoto, joka voi tarttua. myös ihmiseen, minkä jälkeen influenssavirus voi tarttua myös ihmisestä. toiseen.")
26
RNA-VIRUS DNA-VIRUS RETROVIRUS BAKTERIOFAAGI
Viruksia voidaan luokitella niiden genomin perusteella Virusten genomi voi olla: rna:ta joko yksi- tai kaksijuosteisena Kaikkien kaksijuosteisten rna-virusten genomit koostuvat useista erillisistä rna-molekyyleistä (engl. segmented), joskin eräiden yksijuosteistenkin rna-virusten genomit voivat koostua osista. rna-viruksia dna:ta joko yksi- tai kaksijuosteisena. Kaksijuosteisten dna-virusten genomi voi olla joko rengasmaista tai lineaarista (engl. circular vs. linear). dna-viruksia Dna-viruksilla voidaan ajatella olevan etulyöntiasema evoluutiossa rna-viruksiin nähden, sillä perustuuhan dna-virusten hyödyntämä isäntäsolun molekyylibiologien koneisto sekin nimenomaan dna:han. Virusten luokittelussa voi käyttää apuna myös sitä, myös millaisten välivaiheiden kautta valmistuu viruksen genomia vastaavaa m-rna:ta alkuperäisestä viruksen geneettisestä ohjeesta. Kun viruksen rna voi toimia suoraankin m-rna:na, on kyse (+)-juosteviruksista. (-)-juosteinen rna-virus on komplementaarinen vastaavan m-rna:n kanssa. Vasta viruksen rna:sta tehty kopio voi toimia m-rna:na. Retrovirukset ovat rna:ta olevia (+)-juosteviruksia, jotka kuitenkin luokitellaan erityispiirteidensä vuoksi omaan ryhmäänsä. RETROVIRUS BAKTERIOFAAGI
, joskin eräiden yksijuosteistenkin rna-virusten genomit voivat koostua osista. rna-viruksia. dna:ta joko yksi- tai kaksijuosteisena. Kaksijuosteisten dna-virusten genomi voi olla joko rengasmaista tai lineaarista (engl. circular vs. linear). dna-viruksia. Dna-viruksilla voidaan ajatella olevan etulyöntiasema evoluutiossa rna-viruksiin nähden, sillä perustuuhan dna-virusten hyödyntämä isäntäsolun molekyylibiologien koneisto sekin nimenomaan dna:han. Virusten luokittelussa voi käyttää apuna myös sitä, myös millaisten välivaiheiden kautta valmistuu viruksen genomia vastaavaa m-rna:ta alkuperäisestä viruksen geneettisestä ohjeesta. Kun viruksen rna voi toimia suoraankin m-rna:na, on kyse (+)-juosteviruksista. (-)-juosteinen rna-virus on komplementaarinen vastaavan m-rna:n kanssa. Vasta viruksen rna:sta tehty kopio voi toimia m-rna:na. Retrovirukset ovat rna:ta olevia (+)-juosteviruksia, jotka kuitenkin luokitellaan erityispiirteidensä vuoksi omaan ryhmäänsä. RETROVIRUS. BAKTERIOFAAGI.")
27
Bakteriofaagin lisääntyminen
Bakteriofagi on virus, joka loisii bakteerisolussa. Se koostuu joko dna:sta tai rna:sta sekä sitä ympäröivästä proteiinikuoresta. Puolustautuakseen faageilta, bakteerit tuottavat endonukleaasi-proteiineja, jotka pilkkovat dna:ta tietystä emäsjaksosta

28
Dna-viruksen lisääntyminen

29
Retroviruksen lisääntyminen
Viruksista erikoisimpia ovat retrovirukset. Ne ovat genomiltaan yksijuosteista rna:ta, ja ’toimivat’ nurinkurisen käänteisesti verrattuna muuhun eliökuntaan. Retrovirukset kykenevät muuntamaan käänteiskopioinnilla rna-muodossa olevan perimänsä dna-muotoon, vaikka eliöt yleensä kopioivat dna:ta rna-muotoon. Retroviruksen elinkierto. LTR-alueet eli suoratoistot (engl. long terminal repeats) retroviruksen rna:n päissä mahdollistavat käänteiskopioinnin. Retroviruksen koodaama käänteiskopioija-entsyymi ( engl. reverse transcriptase ) tekee ensin viruksen alkuperäinen rna-ohje mallinaan yksijuosteisen dna-molekyylin, joka täydennetään kaksijuosteiseksi dna-molekyyliksi komplementaarisuussääntöjen mukaisesti. Jos käänteiskopioija-entsyymin toiminta pystyttäisiin estämään, olisi se ainakin teoriassa keino retrovirusten aiheuttamien sairauksien, kuten AIDS:in, voittamiseen. Retrovirusten ja ihmisen evolutiivisia suhteita kuvaa se, että yleisimpiä dna-jaksoja ihmisgenomissa on useina satoina kopiona esiintyvä käänteiskopioija-dna. Kaksijuosteinen, 'normaali' dna pystyy toimimaan dna:sta koostuvien liikkuvien jaksojen tavoin ja kiinnittymään isäntägenomiin. Tämän seurauksena kohde-dna:han syntyy suoratoistot aivan kuin liikkuvien jaksojen jäljiltä. Retrovirusten perimää kiinnittyy isäntägenomiin Retroviruksen normaaliin elinkiertoon kuuluu välttämättä vaihe, jolloin se on kiinnittyneenä isäntäsolunsa genomiin dna-muodossa esiviruksena eli proviruksena. Tällöin isäntäsolun geenibiologinen koneisto kohtelee retrovirusperäistä dna:ta kuin omaansa. Samalla syntyy retrovirusperäisä m-rna-molekyylejä retroviruksen proteiinien valmistusta varten sekä pakattaviksi retrovirusjälkeläisten genomeiksi. Retrovirusjälkeläiset ovat diploideja Yksi retrovirusten erikoispiirre on se, että rna-molekyylejä pakataan kuhunkin tekeillä olevaan retrovirusjälkeläiseen kaksi kappaletta, joten jälkeläisistä tulee diploideja. Jos saman solun olisi sattunut infektoimaan kaksi erityypin retrovirusta, voi jälkeläisen genomin rakenne-rna:iksi tulla yksi kumpaakin tyyppiä. Teoriassa ei siis olisi mahdotonta, että esimerkiksi HI-virus ja jokin influenssaa aiheuttava virus joutuisivat samaan soluun. Tällaisen rekombinaation kautta syntyisi siten AIDS:ia aiheuttava influenssavirus, joka leviäisi yhtä tehokkaasti pisaratartuntana ruuhkabusseissa kuin influenssa. Influenssavirukset eivät kuitenkaan ole retroviruksia, vaikka ovatkin (+)-juoste-rna-viruksia, joten käytännössä liian kaukainen sukulaisuus virusten välillä estänee 'tappajainfluenssan' synnyn. Hepatitis B –virus on rappeutunut retrovirus Retrovirukset, joiden genomin omia osia korvautuu tai jää puuttumaan liikaa, eivät enää kykene normaaliin elinkiertoonsa. Tällaisen vajaakykyisen retroviruksen elinkierto on mahdollista vain silloin, kun saman solun infektoi lisäksi toinen virus eli niin sanottu auttajavirus, joka pystyy tuottamaan puuttuvia proteiineja. Esimerkiksi maksatulehdusta aiheuttava hepatitis B -virus on rappeutunut retrovirus, joka on menettänyt kykynsä kiinnittyä genomiin. Retroviruksetkin lisäävät muuntelua kuljettamalla dna-jaksoja Retrovirukset, kuten muutkin virukset, pystyvät elinkiertonsa sivutuotteena lisäämään eliöiden geneettistä monimuotoisuutta kuljettamalla dna:n jaksoja genomista toiseen. Eliöille näistä siirtyvistä jaksoista on kuitenkin todennäköisemmin enemmän haittaa kuin hyötyä. Olisi äärimmäisen harvinainen sattuma, että retroviruksen omaa perimää tai edellisestä isäntäsolusta kaapattua dna:ta kiinnittyisi umpimähkäisesti sellaiseen kohtaan uuden isäntäsolun genomissa, että esimerkiksi syövän kehittymistä edesauttavien soluperäisten geenien toiminta vaimenisi.
 retroviruksen rna:n päissä mahdollistavat käänteiskopioinnin. Retroviruksen koodaama käänteiskopioija-entsyymi ( engl. reverse transcriptase ) tekee ensin viruksen alkuperäinen rna-ohje mallinaan yksijuosteisen dna-molekyylin, joka täydennetään kaksijuosteiseksi dna-molekyyliksi komplementaarisuussääntöjen mukaisesti. Jos käänteiskopioija-entsyymin toiminta pystyttäisiin estämään, olisi se ainakin teoriassa keino retrovirusten aiheuttamien sairauksien, kuten AIDS:in, voittamiseen. Retrovirusten ja ihmisen evolutiivisia suhteita kuvaa se, että yleisimpiä dna-jaksoja ihmisgenomissa on useina satoina kopiona esiintyvä käänteiskopioija-dna. Kaksijuosteinen, normaali dna pystyy toimimaan dna:sta koostuvien liikkuvien jaksojen tavoin ja kiinnittymään isäntägenomiin. Tämän seurauksena kohde-dna:han syntyy suoratoistot aivan kuin liikkuvien jaksojen jäljiltä. Retrovirusten perimää kiinnittyy isäntägenomiin. Retroviruksen normaaliin elinkiertoon kuuluu välttämättä vaihe, jolloin se on kiinnittyneenä isäntäsolunsa genomiin dna-muodossa esiviruksena eli proviruksena. Tällöin isäntäsolun geenibiologinen koneisto kohtelee retrovirusperäistä dna:ta kuin omaansa. Samalla syntyy retrovirusperäisä m-rna-molekyylejä retroviruksen proteiinien valmistusta varten sekä pakattaviksi retrovirusjälkeläisten genomeiksi. Retrovirusjälkeläiset ovat diploideja. Yksi retrovirusten erikoispiirre on se, että rna-molekyylejä pakataan kuhunkin tekeillä olevaan retrovirusjälkeläiseen kaksi kappaletta, joten jälkeläisistä tulee diploideja. Jos saman solun olisi sattunut infektoimaan kaksi erityypin retrovirusta, voi jälkeläisen genomin rakenne-rna:iksi tulla yksi kumpaakin tyyppiä. Teoriassa ei siis olisi mahdotonta, että esimerkiksi HI-virus ja jokin influenssaa aiheuttava virus joutuisivat samaan soluun. Tällaisen rekombinaation kautta syntyisi siten AIDS:ia aiheuttava influenssavirus, joka leviäisi yhtä tehokkaasti pisaratartuntana ruuhkabusseissa kuin influenssa. Influenssavirukset eivät kuitenkaan ole retroviruksia, vaikka ovatkin (+)-juoste-rna-viruksia, joten käytännössä liian kaukainen sukulaisuus virusten välillä estänee tappajainfluenssan synnyn. Hepatitis B –virus on rappeutunut retrovirus. Retrovirukset, joiden genomin omia osia korvautuu tai jää puuttumaan liikaa, eivät enää kykene normaaliin elinkiertoonsa. Tällaisen vajaakykyisen retroviruksen elinkierto on mahdollista vain silloin, kun saman solun infektoi lisäksi toinen virus eli niin sanottu auttajavirus, joka pystyy tuottamaan puuttuvia proteiineja. Esimerkiksi maksatulehdusta aiheuttava hepatitis B -virus on rappeutunut retrovirus, joka on menettänyt kykynsä kiinnittyä genomiin. Retroviruksetkin lisäävät muuntelua kuljettamalla dna-jaksoja. Retrovirukset, kuten muutkin virukset, pystyvät elinkiertonsa sivutuotteena lisäämään eliöiden geneettistä monimuotoisuutta kuljettamalla dna:n jaksoja genomista toiseen. Eliöille näistä siirtyvistä jaksoista on kuitenkin todennäköisemmin enemmän haittaa kuin hyötyä. Olisi äärimmäisen harvinainen sattuma, että retroviruksen omaa perimää tai edellisestä isäntäsolusta kaapattua dna:ta kiinnittyisi umpimähkäisesti sellaiseen kohtaan uuden isäntäsolun genomissa, että esimerkiksi syövän kehittymistä edesauttavien soluperäisten geenien toiminta vaimenisi.")
31
Nimeä viruksen osat

32
Viruksen osat ovat a= vaipan proteiinisauva, B= vaippa, C= proteiinikuori , D= viruksen perimä.

33
Prionit – oikea aminohappojärjestys – väärä proteiini
Prionitaudit aiheutuvat proteiinista Elimistön normaali proteiini muuttuu vääränmuotoiseksi prioniksi Prionin kosketus muuttaa normaalin proteiniinin prioniksi Yhteistä keskushermoston tuhoutuminen Kuva kirjan sivulta 72

34
Proteiinien sekundaarisista rakenteista tavallisimpia ovat alfa-kierre- ja beeta-levyrakenteet. Usein saman yksittäisen polypeptidiketjun väliset vetysidokset saavat aminohappoketjun laskostumaan alfa-kierteiseen muotoon. Beeta-levyksi nimetty levymäinen rakenne taas syntyy tavallisesti silloin, kun kaksi eri, vastakkaissuuntaista polypeptidiketjua liittyvät vetysidoksin toisiinsa. Julkisuudessa suurta kohua herättäneen'hullun lehmän' -taudin (engl. bovine spongiform encephalopathy) aiheuttaa nykytutkimuksen mukaan pelkkä prioniksi nimetty proteiini. Tämä erikoinen taudinaiheuttaja leviää infektoituneiden aivojen ja selkäydinnesteiden välityksellä - yksi ainoa proteiini-infektio saattaa aiheuttaa sairastumisen, joten kannattaa välttää elintarvikkeita, joissa saattaisi olla tätä merkillistä taudinaiheuttajaa. Rituaalista ihmissyöntiä harjoittaneille Uuden Guinealaisen kuru-heimon edustajille ja pässin aivoilla herkutelleille skoteille on käynyt huonosti, kun he samalla olivat tulleet syöneeksi prioneja. Minkään eläinlajin aivojen käyttöä ei nykytietämyksen mukaan voida todellakaan suositella ihmisravinnoksi. Kun ihmissyönti Uudessa Guineassa harvinaistui, myös kuru-tauti eli 'naurava kuolema' hävisi lähes olemattomiin. Carleton Gajdusek, joka tutki kuru-taudin lisäksi samantapaisia Australian alkuasukkaiden tauteja, sai ansioistaan Nobelin palkinnon vuonna 1976 yhdessä Baruch. S. Blumbergin kanssa. 'Hullun lehmän' -tautia aiheuttava prioni on olennaisilta osiltaan beeta-levymäinen proteiini, jonka rakennetta tuskin mitkään tavanomaiset proteiinien hajotuskeinot pystyvät rikkomaan. Näitä tautia aiheuttavia proteiineja vastaavat normaalistikin selkärankaisilla tavattavat, esimerkiksi ihmisen aivoissa tavattavat alfa-kierteiset proteiinit. Prioni -proteiinin keskushermostotauteja aiheuttava versio on siitä merkillinen, että se pystyy muuttamaan haitattomat alfa-kierteiset proteiinit omaa beeta-levymäistä rakennettaan vastaaviksi. Nämä rakenteeltaan muuttuneet proteiinit pystyvät jälleen muuttamaan normaaleja alfa-helikaalisia proteiineja kaltaisekseen. Hullun lehmän taudin tapaisista keskushermostotaudeista tunnetuimpia eläintauteja on lampaan ’scrapie’. Ihmisellä 'hullun lehmän' -tautia vastaavissa perinnöllisissä taudeissa (Gertsman-Straussler-Scheinker - ja Creutzfeldt-Jakob -tauti) yksittäiset aminohappojen muutokset proteiinissa liittyvät taudin syntyyn. Nämä aminohappojen muutokset vaikuttavat ilmeisesti juuri siihen, millaiseksi proteiini muotoutuu sekundaariselta rakenteeltaan.
 aiheuttaa nykytutkimuksen mukaan pelkkä prioniksi nimetty proteiini. Tämä erikoinen taudinaiheuttaja leviää infektoituneiden aivojen ja selkäydinnesteiden välityksellä - yksi ainoa proteiini-infektio saattaa aiheuttaa sairastumisen, joten kannattaa välttää elintarvikkeita, joissa saattaisi olla tätä merkillistä taudinaiheuttajaa. Rituaalista ihmissyöntiä harjoittaneille Uuden Guinealaisen kuru-heimon edustajille ja pässin aivoilla herkutelleille skoteille on käynyt huonosti, kun he samalla olivat tulleet syöneeksi prioneja. Minkään eläinlajin aivojen käyttöä ei nykytietämyksen mukaan voida todellakaan suositella ihmisravinnoksi. Kun ihmissyönti Uudessa Guineassa harvinaistui, myös kuru-tauti eli naurava kuolema hävisi lähes olemattomiin. Carleton Gajdusek, joka tutki kuru-taudin lisäksi samantapaisia Australian alkuasukkaiden tauteja, sai ansioistaan Nobelin palkinnon vuonna 1976 yhdessä Baruch. S. Blumbergin kanssa. Hullun lehmän -tautia aiheuttava prioni on olennaisilta osiltaan beeta-levymäinen proteiini, jonka rakennetta tuskin mitkään tavanomaiset proteiinien hajotuskeinot pystyvät rikkomaan. Näitä tautia aiheuttavia proteiineja vastaavat normaalistikin selkärankaisilla tavattavat, esimerkiksi ihmisen aivoissa tavattavat alfa-kierteiset proteiinit. Prioni -proteiinin keskushermostotauteja aiheuttava versio on siitä merkillinen, että se pystyy muuttamaan haitattomat alfa-kierteiset proteiinit omaa beeta-levymäistä rakennettaan vastaaviksi. Nämä rakenteeltaan muuttuneet proteiinit pystyvät jälleen muuttamaan normaaleja alfa-helikaalisia proteiineja kaltaisekseen. Hullun lehmän taudin tapaisista keskushermostotaudeista tunnetuimpia eläintauteja on lampaan ’scrapie’. Ihmisellä hullun lehmän -tautia vastaavissa perinnöllisissä taudeissa (Gertsman-Straussler-Scheinker - ja Creutzfeldt-Jakob -tauti) yksittäiset aminohappojen muutokset proteiinissa liittyvät taudin syntyyn. Nämä aminohappojen muutokset vaikuttavat ilmeisesti juuri siihen, millaiseksi proteiini muotoutuu sekundaariselta rakenteeltaan.")
35
Ryhmätehtävä Selitä omin sanoin, miksi virukset eivät ole eläviä organismeja. Jos Turkuun julistettaisiin virusepidemia, miten a. terveydenhuollon pitäisi toimia? b. pitäisi toimia, jotta virus ei pääsisi etenemään laajemmalle alueelle? Selitä, miksi antibiootit tehoavat bakteereihin, mutta eivät viruksiin

36
Vastaukset Virukset eivät muodostu soluista. Ne eivät voi tuottaa itsekseen proteiineja. Ne eivät voi käyttää energiaa. Terveydenhuollon pitää valmistautua ottamaan vastaan potilaita, jotka ovat kyseisen viruksen infektoimia. Lisäksi heidän pitää varata riittävästi lääkkeitä, jotka helpottavat taudin oireita. Jos tautiin on ennaltaehkäisevä rokote, rokotuskampanja pitää aloittaa välittömästi. Myös karanteeni mahdollisuus pitää olla olemassa. b. Asiasta on tärkeää tiedottaa, viruksen leviämistie pitää selvittää ja jos mahdollista, se pitää tukkia

37
Antibiootit häiritsevät bakteerisolun toimintoja
Antibiootit häiritsevät bakteerisolun toimintoja. Koska virukset eivät muodostu solusta, häirintä ei toimi niillä.

38
3. Perimän rakenne ja toiminta
1. Perinnöllinen informaatio sijaitsee dna:ssa eli deoksiribonukleiinihapossa 2. Dna sijaitsee kromosomeissa 3. Dna:n kahdentuminen ja solujen jakautuminen siirtää perimätiedon eteenpäin 4. Geenit ohjaavat RNA-molekyylien muodostumista 5. Valkuaisaineet eli proteiinit muodostuvat rna-molekyylien välityksellä 6. Geenien toimintaa säädellään monella eri tasolla 7. Mutaatiot muuttavat perimän viestiä 8. Sukusoluissa tapahtuvat mutaatio siirtyy jälkeläisille ja luonnonvalinnan arvioitavaksi 9. Tehtävät

39
Perinnöllinen informaatio sijaitsee dna:ssa eli deoksiribonukleiinihapossa
Kaksoiskierre Kaksi sokerifosfaattirunkoa Nukleotidi Dna:n rakenneosa Koostuu sokeri-, fosfaatti- ja emäsosasta Emäsparisääntö: A:T, G:C Nukleotidiksi sanotaan nukleiinihappojen, rna:n tai dna:n rakennuspalikkaa, jossa sokeriin on kiinnittynyt jokin emäs sekä fosfori. Emäksistä urasiili on tavallisemmin vain rna:n riboosi-sokeriin kiinnittyneenä, ja tymiini vain dna:n deoksiriboosiin kiinnittyneenä, mutta muut emäkset adeniini, guaniini ja sytosiini voivat esiintyä sekä dna- että rna-molekyyleissä. Nukleosidi on nukleiinihappojen sokeri- ja emäsosista koostuva rakennuspalikka, jossa ei ole fosforia.

40
Dna sisältää emästen lisäksi fosforihappotähteen ja deoksiriboosisokerin.
Emäksiä on kahdenlaisia, sekä puriineja että pyrimidiinejä. Puriiniemäksiä ovat adeniini ja guaniini. Pyrimidiiniemäksiä ovat tymiini ja sytosiini. Adeniinia on aina sama määrä kuin tymiiniä ja sytosiinia yhtä paljon kuin guaniinia. Koska dna-molekyylin kaksi toisiaan vastaavaa juostetta rakentuvat komplementaarisista nukleotideista, myös molekyylin päät ovat komplementaariset. Kummassakin juosteessa on 5'-pää (lue: viisipilkkupää), joka saa nimensä siitä, että uloimman sokerirenkaan viidenteen hiileen kiinnittyy fosforihappo. Juosteen ns. 3'-pää (lue: kolmepilkkupää) tarkoittaa sitä päätä, jossa uloimman sokerin kolmannessa hiilessä on vapaa OH-ryhmä. Tähän vapaaseen 3' OH -ryhmään kiinnittyävät nukleotidit, dna:n rakennuspalikat juosteen pidentyessä. Kaksijuosteisen dna-molekyylin komplementaarisesta rakenteesta seuraa, että yhden juosteen 5'-pää saa parikseen toisen juosteen 3'-pään, ja vastaavasti ensimmäisen juosteen 3'-pää on komplementaarinen toisen juosteen 5'-pään kanssa.
, joka saa nimensä siitä, että uloimman sokerirenkaan viidenteen hiileen kiinnittyy fosforihappo. Juosteen ns. 3 -pää (lue: kolmepilkkupää) tarkoittaa sitä päätä, jossa uloimman sokerin kolmannessa hiilessä on vapaa OH-ryhmä. Tähän vapaaseen 3 OH -ryhmään kiinnittyävät nukleotidit, dna:n rakennuspalikat juosteen pidentyessä. Kaksijuosteisen dna-molekyylin komplementaarisesta rakenteesta seuraa, että yhden juosteen 5 -pää saa parikseen toisen juosteen 3 -pään, ja vastaavasti ensimmäisen juosteen 3 -pää on komplementaarinen toisen juosteen 5 -pään kanssa.")
41
Dna sijaitsee kromosomeissa
Esitumaisilla eliöillä on yksi kromosomi ja rengasmaisia dna-molekyylejä Perimä haploidi Aitotumaisilla dna sijaitsee pääosin tuman kromosomeissa Perimä diploidi tai polyploidi Dna-molekyylit ovat jo perusrakenteeltaan kierteisiä. Aitotumaisten kromosomien muodostuessa molekyylit kiertyvät vielä lisää. Solun genomiin kuuluu useita dna-molekyylejä, eivätkä ne saa sotkeutua keskenään etenkään mitoosissa ja meioosissa solujen jakautuessa. Dna:n kierteisuus saavuttaa huippunsa, solunjakautumisen keskivaiheen (metafaasi) kromosomeissa, jolloin kromosomit ovat valmiina kulkeutumaan jakautuviin tytärsoluihin tumasukkulan avulla. Kaikkein tiivimminkin pakkautuneesta kromosomista voi hapsottaa dna:n edellisiä pakkautumistasoja edustavia lenkkejä. Eräät näistä lenkeistä saattavat kiinnittyä joihinkin tuman sisäkalvon kohtiin. Lenkit purkautuvat helpoiten uloimmista kohdistaan, jotka siten ovat kaikkein alttiimpina replikaatiolle ja transkriptiolle. Silloin dna ei saa olla liian tiivisti pakkautunutta, eivätkä nukleosomit saa olla tiellä. Kromosomirakenteen purkautuminen solukierron välivaihetta, interfaasia varten alkaa sekin juuri lenkkirakenteista.
 kromosomeissa, jolloin kromosomit ovat valmiina kulkeutumaan jakautuviin tytärsoluihin tumasukkulan avulla. Kaikkein tiivimminkin pakkautuneesta kromosomista voi hapsottaa dna:n edellisiä pakkautumistasoja edustavia lenkkejä. Eräät näistä lenkeistä saattavat kiinnittyä joihinkin tuman sisäkalvon kohtiin. Lenkit purkautuvat helpoiten uloimmista kohdistaan, jotka siten ovat kaikkein alttiimpina replikaatiolle ja transkriptiolle. Silloin dna ei saa olla liian tiivisti pakkautunutta, eivätkä nukleosomit saa olla tiellä. Kromosomirakenteen purkautuminen solukierron välivaihetta, interfaasia varten alkaa sekin juuri lenkkirakenteista.")
43
Dna:n pakkautumisen pieneen tilaan mahdollistaa nukleosomirakenne, joka toimii ilmeisesti myös dna:n suojana. Aitotumaisten nukleosomirakenne on osoittautunut evoluutiossa erittäin tärkeäksi, sillä eri lajien nukleosomirakenteeseen vaikuttavat proteiinit ovat eriytyneet tavattoman vähän. Nukleosomirakenne on palautuva (engl. reversible); täytyyhän sen purkautua ennen dna:n kahdentumista sekä silloin, kun geenit toimivat. Toisaalta nukleosomirakenteen täytyy kyetä rakentumaan uudelleen, kun dna:n kahdentumisen tuloksena syntyneet tytär-dna-molekyylit alkavat kiertyä muotoutuakseen tytärkromosomeiksi. Niinpä myös nukleosomirakenteeseen vaikuttavien proteiinien, sekä histonien että muiden proteiinien, täytyy tarvittaessa kaksinkertaistua määrältään. Nukleosomirakenne perustuu juuri histoniproteiineihin. Nämä vahvasti emäksiset proteiinit kiinnittyvät helposti happameen dna-molekyyliin. Histonit ovat ennen kaikkea eukaryooteille eli aitotumallisille eliöille ominaisia dna:n kiertymiseen vaikuttavia proteiineja. Aitotumallisten eliöiden solujen histonien kaukaisia sukulaisproteiineja ovat arkkibakteerien dna:ta kuumuudelta ja happamuudelta suojaavat histonit. Muissakaan bakteerisoluissa ei ole varsinaista nukleosomirakennetta, sillä lenkkirakenteita pursuava bakteerikromosomi on solussa irtonaisena - nukleoidi tarkoittaa bakteerikromosomia tai sen sisältävää aluetta bakteerisolussa. Kun bakteerisolu jakautuu, ei tarvita eukaryoottien tumasukkulan tapaisia keinoja kromosomien tasapuoliseen jakoon tytärsolujen kesken. Sen sijaan tytärkromosomit takertuvat joistakin lenkeistään kiinni sopiviin kohtiin bakteerisolun kalvon sisäpintaan. Bakteerisolun jakautuessa kumpaakin tytärsoluun jää siten yleensä yhtä monta kromosomia.
; täytyyhän sen purkautua ennen dna:n kahdentumista sekä silloin, kun geenit toimivat. Toisaalta nukleosomirakenteen täytyy kyetä rakentumaan uudelleen, kun dna:n kahdentumisen tuloksena syntyneet tytär-dna-molekyylit alkavat kiertyä muotoutuakseen tytärkromosomeiksi. Niinpä myös nukleosomirakenteeseen vaikuttavien proteiinien, sekä histonien että muiden proteiinien, täytyy tarvittaessa kaksinkertaistua määrältään. Nukleosomirakenne perustuu juuri histoniproteiineihin. Nämä vahvasti emäksiset proteiinit kiinnittyvät helposti happameen dna-molekyyliin. Histonit ovat ennen kaikkea eukaryooteille eli aitotumallisille eliöille ominaisia dna:n kiertymiseen vaikuttavia proteiineja. Aitotumallisten eliöiden solujen histonien kaukaisia sukulaisproteiineja ovat arkkibakteerien dna:ta kuumuudelta ja happamuudelta suojaavat histonit. Muissakaan bakteerisoluissa ei ole varsinaista nukleosomirakennetta, sillä lenkkirakenteita pursuava bakteerikromosomi on solussa irtonaisena - nukleoidi tarkoittaa bakteerikromosomia tai sen sisältävää aluetta bakteerisolussa. Kun bakteerisolu jakautuu, ei tarvita eukaryoottien tumasukkulan tapaisia keinoja kromosomien tasapuoliseen jakoon tytärsolujen kesken. Sen sijaan tytärkromosomit takertuvat joistakin lenkeistään kiinni sopiviin kohtiin bakteerisolun kalvon sisäpintaan. Bakteerisolun jakautuessa kumpaakin tytärsoluun jää siten yleensä yhtä monta kromosomia.")
44
Dna:n kahdentuminen ja solujen jakautuminen siirtää perimätiedon eteenpäin
Kahdentuminen tapahtuu ennen solun jakautumista Kumpikin juoste toimii uuden juosteen mallina Replikaatio Emäsparien väliset vetysidokset katkeavat Etenee aloituskohdasta kumpaankin suuntaan Dna-polymeraasientsyymi rakentajana Vastinjuosteet kopioituvat vastakkaisiin suuntiin Jotta periytyminen ylipäätään olisi mahdollista, täytyy dna:ssa sijaitsevien geenien kahdentua ennen jokaista solunjakautumista. Dna:n kahdentumiskykyyn perustuu sekä evoluution alkuhämärissä menestyneiden kahdentumiskykyisten molekyylien runsastuminen että nykyisten solujen olemassaolo. Koska ilman soluja ei olisi eliöitäkään, voidaan dna:n kahdentumista pitää sekä bakteerin että ihmisen olemassaolon yhtenä olennaisimpana edellytyksenä. Tästä vastaavan solujen biologisen järjestelmän mahdollisimman virheetön toiminta on tae olemassaolomme jatkuvuudesta sekä somaattisten solujen muodostaman elimistön soluvaltion että sukusoluista alkunsa saavien tulevien sukupolvien suhteen. Dna:n kahdentumisen jälkeen tapahtuva dna:n kiertyminen kromosomeiksi on puolestaan edellytys sekä kromosomien vähennysjakautumiselle meioosissa että kromosomien mitoottisille jaoille. Dna:n kahdentuminen avaakin reitin, joka johtaa solun jakautumiseen ja tytärsolujen syntyyn. Dna:n kahdentumisen ajoitus on tarkasti säädeltyä. Dna:n kahdentumisen, kiertymisen kromosomeiksi ja tumasukkulan avulla tapahtuvien kromosomien liikkeiden täytyy olla synkronisoituna solun kahdentumiseen ja jakautumiseen. Tätä solusykliksi kutsuttua solun 'elinkiertoa' säätelevät monet geenit tuottamiensa proteiinien välityksellä. Tärkeimpiä solusykliä sääteleviä proteiineja ovat muita proteiineja fosforyloivat proteiinit. Syöpätutkimuksessa keskitytäänkin solusyklin poikkeavuuksiin, joiden ymmärtämistä varten täytyy tuntea normaali solusykli mahdollisimman hyvin Varsinaisessa dna:n kahdentumisessa dna-kaksoiskierre avautuu ja juosteet irtoavat toisistaan ikään kuin vetoketjua avatessa. Kummankin juosteen viereen rakentuu komplementaarinen uusi juoste nukleotideista, dna:n rakennuspalikoista. Näin syntyvät puoliksi uudet molekyylit, jotka kummatkin koostuvat sekä uudesta että vanhasta juosteesta - dna:n kahdentuminen tapahtuu siis semikonservatiivisesti. Niinpä ei teoriassa ole mahdotonta, että soluissasi sattuisi olemaan ikivanha nukleotidi, joka on peräisin Afrikassa eläneeltä esivanhemmaltamme. Tai jokin evoluution alkuhämärästä peräisin olevan nukleotidiparin toinen osapuoli olisi päätynyt soluihisi ja toinen osapuoli olisi kulkeutunut koirasi tai kissasi soluihin. Dna:n kahdentumisen päästyä vauhtiin, on tärkein entsyymi uutta juostetta valmistava dna-polymeraasi. Dna-polymeraaseja on useaa eri tyyppiä, ja jokainen niistä on erikoistunut tietynlaisiin tehtäviin. Muun muassa dna:n kahdentumisessa syntyvien virheiden korjaamiseen tarvitaan juuri tähän tehtävään erikoistuneet dna-polymeraasit. Aitotumaisilla dna-polymeraaseja on viittä eri tyyppiä, E. coli-bakteerilla kolme erilaista (dna-polymeraasit I, II ja III). Dna:n kahdentuminen onkin tavattoman tarkkaa. Bakteerien dna:n kahdentuessa syntyy keskimäärin yksi virhe genomia kohden 1000 replikaatiossa. Jos tekstinkäsittelyssä yksi merkki vastaisi yhtä emästä, pystyisi eräs bakteerien dna-polymeraaseista, dna-polymeraasi III, kopioimaan viisi miljoonaa sivua tekstiä virheettömästi. Esimerkiksi biologian oppikirjan tekstiä tämä polymeraasi kykenisi kopioimaan yhtäjaksoisesti vähintään noin kirjasta, ennen ensimmäistä tilastollista kopioimisvirhettä. Etenkin E. Coli –bakteerilla on tutkittu replikaation tarkkuutta. Dna:n kahdentumisen virheettömyys johtuu ennen kaikkea dna-polymeraasi III:n 3' -5' -eksonukleaasiaktiivisuudesta. Tämä merkitsee sitä, että entsyymi tarkistaa, onko komplementaarinen nukleotidi liittymässä syntyvän juosteen jatkoksi. 3' - 5' eksonukleaasiaktiivisuuden avulla dna-polymeraasi III voi poistaa epäkomplementaarisen emäksen sisältämän nukleotidin yksi kerrallaan ennen sen kiinnittymistä juosteeseen. Kaikilla E. colin dna-polymeraaseilla (I, II, III) on 3'-5 -eksonukleaasiaktiivisuus. Ainoastaan virheitä korjaavalla dna-polymeraasi I:llä on myös 5'-3' - eksonukleaasiaktiivisuus. Niinpä tämä entsyymi pystyy nukleotidin kiinnittymisen jälkeenkin tarkistamaan komplementaarisuuden. Mikäli komplementaarisuussääntöjen vastainen emäs on liitetty syntyvään juosteeseen, dna-polymeraasi I voi irrottaa sen. dna-polymeraasi I voi irrottaa myös pidempiä jaksoja, koostuivat ne sitten riboosi- tai deoksiriboosinukleotideistä. Kaikki tunnetut dna-polymeraasit, jotka valmistavat uutta juostetta, liittävät uuden nukleotidin entisen perään 5' - 3' -suunnassa. Vain tällöin on nukleotidin sokerin kolmannessa hiilessä vapaa OH-ryhmä, johon uusi nukleotidi liitetään fosforiosastaan. Replikaation aloituksessa tarvittaviin proteiineihin kuuluu (dna G-geenin tuottama) primaasi-proteiini, joka pystyy valmistamaan lyhyitä jaksoja rna:ta. Näitä rna-alukkeita tarvitaan dna:ta valmistavaa entsyymiä, dna-polymeraasia varten. Dna-polymeraasi ei pysty aloittamaan dna:n valmistusta muuten, kuin jatkamalla nukleotidien lisäämistä jo olemassaolevaan rna-alukkeeseen. On aivan välttämätöntä, että vähintään kaksi ensimmäistä alukkeen nukleotidia ovat rna:ta. Primaasi-proteiini pystyy kyllä (dna B:n avustamana) valmistamaan alukkeen joko deoksiriboosia tai riboosia sisältävistä nukleotideista. Koska solussa on yleensä ylimäärin riboosia sisältäviä nukleotideja verrattuna deoksiriboosia sisältäviin nukleotideihin, on syntyvä aluke yleensä rna:ta muutenkin kuin kahden ensimmäisen nukleotidin osalta. Primosomi, aloitusproteiinien kasauma tunnistaa hiuspinni-lenkin, jonka viereen alkaa rakentua rna-aluke. Liikkuessaan pitkin dna:ta tämä proteiinien kasauma tekee alukkeita sinne tänne. Lineaaristen kromosomien päiden replikaatio tapahtuu telomeraasin avulla Aitotumaisten eliöiden lineaaristen kromosomien päiden replikaation seurauksena kromosomit lyhenisivät joka replikaatiokierroksella koska osista koostuva laahaava juoste ei voisi enää pidentyä, kun viimeisen osan alukkeelle ei enää löytyisi vastaavaa kohtaa vanhassa juosteessa. Vanhan juosteen 3’-päähän kiinnittyvä telomeraasi-entsyymi pystyy rna-alukkeensa avulla replikoimaan myös kromosomien päät. Kromosomin päiden replikaatiopulma voidaan kiertää myös muilla keinoin. Muun muassa adenoviruksen, rna-polioviruksen ja joidenkin faagien käyttämä replikaatiokoneisto pääsee toisen mallijuosteen alkuun sen 5'-päähän kiinnittyneen proteiinin avulla. Tämä proteiini sisältää OH-ryhmän, joka vastaa tavanomaista 3'-pään vapaata OH-ryhmää.
. Dna:n kahdentuminen onkin tavattoman tarkkaa. Bakteerien dna:n kahdentuessa syntyy keskimäärin yksi virhe genomia kohden 1000 replikaatiossa. Jos tekstinkäsittelyssä yksi merkki vastaisi yhtä emästä, pystyisi eräs bakteerien dna-polymeraaseista, dna-polymeraasi III, kopioimaan viisi miljoonaa sivua tekstiä virheettömästi. Esimerkiksi biologian oppikirjan tekstiä tämä polymeraasi kykenisi kopioimaan yhtäjaksoisesti vähintään noin kirjasta, ennen ensimmäistä tilastollista kopioimisvirhettä. Etenkin E. Coli –bakteerilla on tutkittu replikaation tarkkuutta. Dna:n kahdentumisen virheettömyys johtuu ennen kaikkea dna-polymeraasi III:n eksonukleaasiaktiivisuudesta. Tämä merkitsee sitä, että entsyymi tarkistaa, onko komplementaarinen nukleotidi liittymässä syntyvän juosteen jatkoksi eksonukleaasiaktiivisuuden avulla dna-polymeraasi III voi poistaa epäkomplementaarisen emäksen sisältämän nukleotidin yksi kerrallaan ennen sen kiinnittymistä juosteeseen. Kaikilla E. colin dna-polymeraaseilla (I, II, III) on eksonukleaasiaktiivisuus. Ainoastaan virheitä korjaavalla dna-polymeraasi I:llä on myös eksonukleaasiaktiivisuus. Niinpä tämä entsyymi pystyy nukleotidin kiinnittymisen jälkeenkin tarkistamaan komplementaarisuuden. Mikäli komplementaarisuussääntöjen vastainen emäs on liitetty syntyvään juosteeseen, dna-polymeraasi I voi irrottaa sen. dna-polymeraasi I voi irrottaa myös pidempiä jaksoja, koostuivat ne sitten riboosi- tai deoksiriboosinukleotideistä. Kaikki tunnetut dna-polymeraasit, jotka valmistavat uutta juostetta, liittävät uuden nukleotidin entisen perään suunnassa. Vain tällöin on nukleotidin sokerin kolmannessa hiilessä vapaa OH-ryhmä, johon uusi nukleotidi liitetään fosforiosastaan. Replikaation aloituksessa tarvittaviin proteiineihin kuuluu (dna G-geenin tuottama) primaasi-proteiini, joka pystyy valmistamaan lyhyitä jaksoja rna:ta. Näitä rna-alukkeita tarvitaan dna:ta valmistavaa entsyymiä, dna-polymeraasia varten. Dna-polymeraasi ei pysty aloittamaan dna:n valmistusta muuten, kuin jatkamalla nukleotidien lisäämistä jo olemassaolevaan rna-alukkeeseen. On aivan välttämätöntä, että vähintään kaksi ensimmäistä alukkeen nukleotidia ovat rna:ta. Primaasi-proteiini pystyy kyllä (dna B:n avustamana) valmistamaan alukkeen joko deoksiriboosia tai riboosia sisältävistä nukleotideista. Koska solussa on yleensä ylimäärin riboosia sisältäviä nukleotideja verrattuna deoksiriboosia sisältäviin nukleotideihin, on syntyvä aluke yleensä rna:ta muutenkin kuin kahden ensimmäisen nukleotidin osalta. Primosomi, aloitusproteiinien kasauma tunnistaa hiuspinni-lenkin, jonka viereen alkaa rakentua rna-aluke. Liikkuessaan pitkin dna:ta tämä proteiinien kasauma tekee alukkeita sinne tänne. Lineaaristen kromosomien päiden replikaatio tapahtuu telomeraasin avulla. Aitotumaisten eliöiden lineaaristen kromosomien päiden replikaation seurauksena kromosomit lyhenisivät joka replikaatiokierroksella koska osista koostuva laahaava juoste ei voisi enää pidentyä, kun viimeisen osan alukkeelle ei enää löytyisi vastaavaa kohtaa vanhassa juosteessa. Vanhan juosteen 3’-päähän kiinnittyvä telomeraasi-entsyymi pystyy rna-alukkeensa avulla replikoimaan myös kromosomien päät. Kromosomin päiden replikaatiopulma voidaan kiertää myös muilla keinoin. Muun muassa adenoviruksen, rna-polioviruksen ja joidenkin faagien käyttämä replikaatiokoneisto pääsee toisen mallijuosteen alkuun sen 5 -päähän kiinnittyneen proteiinin avulla. Tämä proteiini sisältää OH-ryhmän, joka vastaa tavanomaista 3 -pään vapaata OH-ryhmää.")
45
Geenit ohjaavat RNA-molekyylien muodostumista
Geenissä säätelyalue ja rna-molekyyliksi kopioitava alue Eksoni sisältää tiedon proteiinisynteesistä Introni (eksonien välissä) eivät sisällä tietoa poistetaan ennen esiaste-rna:sta ennen proteiinisynteesiä Toistojaksot tarjoavat materiaalia dna-sormenjälkiä varten Säätelyalueet ohjaavat geenien toimintaa: Promoottorialueeseen kiinnittyy dna-polymeraasi Käynnistävät transkription Transkriptiofaktorit kiinnittyvät säätelyalueisiin ja tehostajajaksoihin säätelevät kaksoiskierteen aukeamista ja rna-polymeraasin toimintaa Eri geenit toimivat eri kudoksissa eri kehitysvaiheiden aikana. Taloudenpitogeeneiksi sanotaan geenejä, jotka tuottavat soluille välttämättämiä proteiineja, ja jotka toimivat tasaisesti lähes kaiken aikaa. Ylellisyysgeeneiksi sanotaan geenejä, joiden tuotteet eivät ole soluille välttämättömiä, mutta joista tietyissä tilanteissa saattaa olla hyötyä. Esimerkiksi bakteerien antibioottiresistenssigeenejä voi pitää ylellisyysgeeneinä, kun taas ihmisen hemoglobiini –proteiinin esiasteita täytyy elimistössä valmistua jatkuvasti. Geenien toiminta eli ilmeneminen onkin siksi tarkasti säädeltyä. Geenit ilmenevät, jos niiden transkriptio lähtee käyntiin. Transkriptiossa valmistuu m-rna-molekyylejä, jonka koodin mukaan translaatiossa liitetään aminohapot peptidiketjuksi. Transkriptio on tärkeä vaihe geenisäätelyssä – sitä voikin pitää geenien toiminnan pullonkaulana Yksinkertaisimmillaan eukaryootin dna:ssa on varsinaisen proteiinia koodaavan jakson edeltäjänä säätelyalue. Säätelyalueen eri kohtiin sitoutuu: käynnistäjäproteiineja m-rna:n (engl. messenger) eli lähetti-rna:n syntyyn tarvittava rna-polymeraasi promoottoriksi sanotulle alueelle Sekä eukaryooteilla että prokaryooteilla on transkription aloituskohdan lähellä jaksoja, joissa on oltava tietynlainen emäsjärjestys (TATA/TATAAT). On myös mahdollista, että säätelyalueita on useita ja ne voivat sijaita kaukanakin varsinaisesta koodaavasta jaksosta. Geenisäätelyyn osallistuvia proteiineja voi myös olla lukuisia – joissain tapauksissa jopa satoja. Nykytutkimuksessa selvitetään juuri näitä proteiinien välisiä vuorovaikutuksia. Kaiken kaikkiaan geenien toiminnan säätely voi perustua: proteiineihin dna-jaksoihin rna-molekyyleihin Säätely voi olla positiivista tai negatiivista. Negatiivisessa säätelyssä estäjäproteiini(t) on sitoutuneena säätelyalueelle, eikä transkriptio lähde käyntiin ennen kuin estäjä irtoaa. Positiivisessa säätelyssä tarvitaan proteiini, joka sitoutuu säätelyalueelle, jotta transkriptio lähtee käyntiin. Transkriptiossa dna-molekyylin mallin mukaan valmistuu rna-molekyyli, m-rna. m-rna-lyhenne juontuukin englannin kielen messenger-sanasta, jonka merkitys on 'välittäjä', 'viestinviejä'. M-rna:ta tarvitaan, jotta tieto geenihin koodatusta proteiinien valmistusohjeesta voidaan viedä solun tumasta solulimaan, missä proteiinienvalmistuskoneisto toimii. Dna-molekyyli on kaksijuosteinen, mutta m-rna-molekyyli, kuten rna yleensäkin on yksijuosteista. Niinpä m-rna-molekyyli rakentuu dna:n toisen juosteen viereen, jonka kanssa se on komplementaarinen. Tätä dna:n juostetta kutsutaan ei-koodaavaksi juosteeksi, sillä koodaava juoste on m-rna:n kanssa identtinen juoste (lukuunottamatta tymiini-urasiili-eroa). Juuri koodaavan juosteen (engl. sense) geneettinen informaatio siirtyy transkriptiossa eteen päin, tosin käännettynä -transkriptoituna - rna-muotoon. Ennen digikuvauksen valtakautta valokuvauksessakin sai positiivikuvasta toisen positiivikuvan vain negatiivivälivaiheen kautta.
 eivät sisällä tietoa poistetaan ennen esiaste-rna:sta ennen proteiinisynteesiä. Toistojaksot tarjoavat materiaalia dna-sormenjälkiä varten. Säätelyalueet ohjaavat geenien toimintaa: Promoottorialueeseen kiinnittyy dna-polymeraasi. Käynnistävät transkription. Transkriptiofaktorit kiinnittyvät säätelyalueisiin ja tehostajajaksoihin säätelevät kaksoiskierteen aukeamista ja rna-polymeraasin toimintaa. Eri geenit toimivat eri kudoksissa eri kehitysvaiheiden aikana. Taloudenpitogeeneiksi sanotaan geenejä, jotka tuottavat soluille välttämättämiä proteiineja, ja jotka toimivat tasaisesti lähes kaiken aikaa. Ylellisyysgeeneiksi sanotaan geenejä, joiden tuotteet eivät ole soluille välttämättömiä, mutta joista tietyissä tilanteissa saattaa olla hyötyä. Esimerkiksi bakteerien antibioottiresistenssigeenejä voi pitää ylellisyysgeeneinä, kun taas ihmisen hemoglobiini –proteiinin esiasteita täytyy elimistössä valmistua jatkuvasti. Geenien toiminta eli ilmeneminen onkin siksi tarkasti säädeltyä. Geenit ilmenevät, jos niiden transkriptio lähtee käyntiin. Transkriptiossa valmistuu m-rna-molekyylejä, jonka koodin mukaan translaatiossa liitetään aminohapot peptidiketjuksi. Transkriptio on tärkeä vaihe geenisäätelyssä – sitä voikin pitää geenien toiminnan pullonkaulana. Yksinkertaisimmillaan eukaryootin dna:ssa on varsinaisen proteiinia koodaavan jakson edeltäjänä säätelyalue. Säätelyalueen eri kohtiin sitoutuu: käynnistäjäproteiineja. m-rna:n (engl. messenger) eli lähetti-rna:n syntyyn tarvittava rna-polymeraasi promoottoriksi sanotulle alueelle. Sekä eukaryooteilla että prokaryooteilla on transkription aloituskohdan lähellä jaksoja, joissa on oltava tietynlainen emäsjärjestys (TATA/TATAAT). On myös mahdollista, että säätelyalueita on useita ja ne voivat sijaita kaukanakin varsinaisesta koodaavasta jaksosta. Geenisäätelyyn osallistuvia proteiineja voi myös olla lukuisia – joissain tapauksissa jopa satoja. Nykytutkimuksessa selvitetään juuri näitä proteiinien välisiä vuorovaikutuksia. Kaiken kaikkiaan geenien toiminnan säätely voi perustua: proteiineihin. dna-jaksoihin. rna-molekyyleihin. Säätely voi olla positiivista tai negatiivista. Negatiivisessa säätelyssä estäjäproteiini(t) on sitoutuneena säätelyalueelle, eikä transkriptio lähde käyntiin ennen kuin estäjä irtoaa. Positiivisessa säätelyssä tarvitaan proteiini, joka sitoutuu säätelyalueelle, jotta transkriptio lähtee käyntiin. Transkriptiossa dna-molekyylin mallin mukaan valmistuu rna-molekyyli, m-rna. m-rna-lyhenne juontuukin englannin kielen messenger-sanasta, jonka merkitys on välittäjä , viestinviejä . M-rna:ta tarvitaan, jotta tieto geenihin koodatusta proteiinien valmistusohjeesta voidaan viedä solun tumasta solulimaan, missä proteiinienvalmistuskoneisto toimii. Dna-molekyyli on kaksijuosteinen, mutta m-rna-molekyyli, kuten rna yleensäkin on yksijuosteista. Niinpä m-rna-molekyyli rakentuu dna:n toisen juosteen viereen, jonka kanssa se on komplementaarinen. Tätä dna:n juostetta kutsutaan ei-koodaavaksi juosteeksi, sillä koodaava juoste on m-rna:n kanssa identtinen juoste (lukuunottamatta tymiini-urasiili-eroa). Juuri koodaavan juosteen (engl. sense) geneettinen informaatio siirtyy transkriptiossa eteen päin, tosin käännettynä -transkriptoituna - rna-muotoon. Ennen digikuvauksen valtakautta valokuvauksessakin sai positiivikuvasta toisen positiivikuvan vain negatiivivälivaiheen kautta.")
47
Valkuaisaineet eli proteiinit muodostuvat rna-molekyylien välityksellä
Proteiinisynteesi alkaa, kun lähetti-rna kiinnittyy ribosomiin solulimassa Siirtäjä-rna:t kuljettavat aminohappoja ribosomille. Lähetti-rna:n jokaista erilaista kodonia varten on oma siirtäjä-rna-molekyylinsä. Aloituskolmikko aina ATG (metioniini) mallijuoste TAC lähetti-rna AUG siirtäjä-rna UAC Antikodoni eli siirtäjä-rna:n tunnistuskolmikko pariutuu lähetti-rna:ssa emäsparisäännön mukaan Lopetuskodonit ovat ATT, ATC ja ACT Translaatiossa dna:ssa oleva geneettinen informaatio, proteiinin valmistusohje, toteutuu m-rna:n välityksellä. Myös tämä vaihe, kuten suurin osa geenibiologista koneistoa, tunnetaan parhaiten prokaryooteilta. M-rna välittää tiedon siitä, mihin järjestykseen aminohapot liitetään valmistuvassa polypeptidissä. M-rna:n luenta ribosomeilla ohjaa aminohapot oikeaan järjestykseen siirtäjä-rna-molekyylien välityksellä. Geneettinen koodi merkitseekin sitä, että kolme perättäistä emästä dna:ssa eli kodoni vastaa yhtä aminohappoa proteiinissa. Koska nukleotideissa voi olla mikä tahansa neljästä emäksestä, on periaatteessa 64 mahdollista (43) kolmen emäksen koodia. Aminohappoja on kuitenkin vain 20, joten useampi kodoni voi vastata yhtä aminohappoa, vaikka kolme kodonia toimiikin lopetuskodoneina. Niitä aminohappoja, joilla on eniten vaihtoehtoisia kodoneja, esiintyy myös eniten proteiineissa. Geneettisen koodin degeneraatio eli rappeutuminen tarkoittaa, että kodonit voivat erota etenkin kolmannen emäksen suhteen, mutta tästä huolimatta ne ohjaavat samaa laatua olevan aminohapon valmistuvaan proteiiniin. Kolmas emäs saattaa näet olla täysin merkityksetön, tai sitten se voi vaihdella ollen ylipäätään joko puriini tai pyrimidiini. Metioniini ja tryptofaani ovat siitä epätyypillisiä aminohappoja, että kummallekin on vain yksi tietty kodoni - kodonin kolmannessa emäksessäkään ei ole vaihtelua. Geneettinen koodi on peruspiirteiltään samanlainen kaikissa eliöissä, joskin joidenkin kodonien suhteen eri eliöt ovat kehittyneet omaan suuntaansa. Vuonna 1983 löydettiin ensimmäiset geneettisen koodin poikkeamat mitokondrioista. Mitokondrioissa ne kodonit, jotka muilla eliöillä koodaavat aminohappoja toimivatkin lopetuskodoneina ja tryptofaanille ja metioniinille on kummallekin kaksi kodonia. Tämän jälkeen huomattiin esimerkiksi siimaeliöiden ja Mycoplasma-bakteerien UAA- ja UAG -kodonien koodaavan glutamiinia, vaikka ne muilla eliöillä toimivat lopetuskodoneina. Avoin lukukehys viittaa tuntemattomaan geeniin Geneettinen koodi keksittiin vuonna 1961, jolloin havaittiin poly-U-jaksojen m-rna:ssa johtavan peräkkäisiin fenyylialaniini -nimisten aminohappojen kertymiin proteiinissa (Nirenberg). Sittemmin keinotekoisten nukleotidijaksojen avulla emäsjärjestystä systemaattisesti vaihdellen ratkaistiin lopulta se, mikä tietty kodoni vastaa mitäkin aminohappoa. Avoin lukukehys merkitseekin dna-jaksoa, jossa aloitus- ja lopetuskodonien väli sisältää aminohappoja koodaavia kolmen emäksen yhdistelmiä, eikä mitä tahansa merkityksettömiä emäsjonoja. Käytännössä avoin lukukehys viittaa tunnistamattomaan geeniin, josta halutaan selkoa. Esimerkiksi ihmisgenomin kartoituksen yhtenä hankaluutena on se, että avoimet lukukehykset saattavat sijaita limittäin. Virukset ja faagit puolestaan vaihtelevat lukukehyksen vaiheistusta, mikä monimutkaistaa sekvenointimenetelmillä selvitetyn emäsjärjestyksen tulkintaa suhteessa mahdollisiin aminohappojärjestyksiin.
 mallijuoste TAC. lähetti-rna AUG. siirtäjä-rna UAC. Antikodoni eli siirtäjä-rna:n tunnistuskolmikko pariutuu lähetti-rna:ssa emäsparisäännön mukaan. Lopetuskodonit ovat ATT, ATC ja ACT. Translaatiossa dna:ssa oleva geneettinen informaatio, proteiinin valmistusohje, toteutuu m-rna:n välityksellä. Myös tämä vaihe, kuten suurin osa geenibiologista koneistoa, tunnetaan parhaiten prokaryooteilta. M-rna välittää tiedon siitä, mihin järjestykseen aminohapot liitetään valmistuvassa polypeptidissä. M-rna:n luenta ribosomeilla ohjaa aminohapot oikeaan järjestykseen siirtäjä-rna-molekyylien välityksellä. Geneettinen koodi merkitseekin sitä, että kolme perättäistä emästä dna:ssa eli kodoni vastaa yhtä aminohappoa proteiinissa. Koska nukleotideissa voi olla mikä tahansa neljästä emäksestä, on periaatteessa 64 mahdollista (43) kolmen emäksen koodia. Aminohappoja on kuitenkin vain 20, joten useampi kodoni voi vastata yhtä aminohappoa, vaikka kolme kodonia toimiikin lopetuskodoneina. Niitä aminohappoja, joilla on eniten vaihtoehtoisia kodoneja, esiintyy myös eniten proteiineissa. Geneettisen koodin degeneraatio eli rappeutuminen tarkoittaa, että kodonit voivat erota etenkin kolmannen emäksen suhteen, mutta tästä huolimatta ne ohjaavat samaa laatua olevan aminohapon valmistuvaan proteiiniin. Kolmas emäs saattaa näet olla täysin merkityksetön, tai sitten se voi vaihdella ollen ylipäätään joko puriini tai pyrimidiini. Metioniini ja tryptofaani ovat siitä epätyypillisiä aminohappoja, että kummallekin on vain yksi tietty kodoni - kodonin kolmannessa emäksessäkään ei ole vaihtelua. Geneettinen koodi on peruspiirteiltään samanlainen kaikissa eliöissä, joskin joidenkin kodonien suhteen eri eliöt ovat kehittyneet omaan suuntaansa. Vuonna 1983 löydettiin ensimmäiset geneettisen koodin poikkeamat mitokondrioista. Mitokondrioissa ne kodonit, jotka muilla eliöillä koodaavat aminohappoja toimivatkin lopetuskodoneina ja tryptofaanille ja metioniinille on kummallekin kaksi kodonia. Tämän jälkeen huomattiin esimerkiksi siimaeliöiden ja Mycoplasma-bakteerien UAA- ja UAG -kodonien koodaavan glutamiinia, vaikka ne muilla eliöillä toimivat lopetuskodoneina. Avoin lukukehys viittaa tuntemattomaan geeniin. Geneettinen koodi keksittiin vuonna 1961, jolloin havaittiin poly-U-jaksojen m-rna:ssa johtavan peräkkäisiin fenyylialaniini -nimisten aminohappojen kertymiin proteiinissa (Nirenberg). Sittemmin keinotekoisten nukleotidijaksojen avulla emäsjärjestystä systemaattisesti vaihdellen ratkaistiin lopulta se, mikä tietty kodoni vastaa mitäkin aminohappoa. Avoin lukukehys merkitseekin dna-jaksoa, jossa aloitus- ja lopetuskodonien väli sisältää aminohappoja koodaavia kolmen emäksen yhdistelmiä, eikä mitä tahansa merkityksettömiä emäsjonoja. Käytännössä avoin lukukehys viittaa tunnistamattomaan geeniin, josta halutaan selkoa. Esimerkiksi ihmisgenomin kartoituksen yhtenä hankaluutena on se, että avoimet lukukehykset saattavat sijaita limittäin. Virukset ja faagit puolestaan vaihtelevat lukukehyksen vaiheistusta, mikä monimutkaistaa sekvenointimenetelmillä selvitetyn emäsjärjestyksen tulkintaa suhteessa mahdollisiin aminohappojärjestyksiin.")
49
Geenien toimintaa säädellään monella eri tasolla
Tuottajageenit Ohjaavat solun rakenneproteiinien ja entsyymien muodostumista Reagoivat säätelygeenien tuottamiin säätelytekijöihin Säätelygeenit Säätelevät muiden geenien toimintaa mm. proteiinien avulla Säätelevät: geenien transkriptiota, lähetti-rna:n muokkausta, kulkeutumista solulimaan, hajotusta sekä proteiinisynteesiä. Silmukointi: esiaste-rna:n vaihtoehtoinen muokkaus yhdestä geenistä voidaan tuottaa kymmeniä erilaisia proteiineja Kuva kirjan sivultaMutaatioiden perusteella on pääteltävissä, millainen säätelyjärjestelmä on kyseessä. Esimerkiksi laktoosioperoniin voi kohdistua erilaisia mutaatioita, joilla on erilaisia vaikutuksia. Varsinainen käynnistäjäalue voi muteerata niin, ettei rna-polymeraasi kykene sitoutumaan siihen tavalliseen tapaansa - silloin laktoosioperonin geenit eivät ilmene koskaan. Myös repressorin sitoutumiskohta dna:ssa voi muteerata, jolloin repressori ei voi kiinnittyä siihen - silloin operonin geenit ovat aina toiminnassa. Mikäli mutaatio kohdistuu repressoria koodaavaan erilliseen säätelygeeniin, ei virheellinen repressori kykene irtoamaan käynnistäjästä laktoosin indusoimanakaan. Säätelijägeenit ovat tärkeitä geenejä, sillä ne vaikuttavat koodamiensa proteiinien välityksellä muiden geenien ilmenemiseen. Siten niiden vaikutus kohdistuu laajaan geenijoukkoon. Rakennegeenit ovat varsinaisia proteiineja koodaavia geenejä, joiden ilmenemiseen säätelevät proteiinit vaikuttavat. Välttämättömät taloudenpitogeenit ovat juuri rakennegeenejä, joskin toisinaan myös pelkkää rna:ta tuottavat geenit luokitellaan rakennegeeneihin. Mikäli yksittäiseen säätelygeeniin kohdistuu mutaatio, saattaa useiden eri rakennegeenien koodaamien proteiinien määrä muuttua niiden tavanomaiseen tasoon verrattuna. Transkription lopetukseen vaikuttavat myös useat eri proteiinit, jotka ovat tyypillisiä eri bakteereille ja faageille. Näistä proteiineistä toiset ovat välttämättömiä, jotta transkriptio loppuu, toiset taas aiheuttavat transkription jatkumisen geenin ohi. Mikäli transkriptio etenee yhdestä geenistä seuraavaan, näin syntyvään m-rna-molekyyliin sisältyy usean geenin koodi. Etenkin faageilla esiintyy sellaisia transkription lopetusjaksoja, joihin sitoutuva rho-proteiini on välttämätön transkription pysähtymiselle.

50
Paitsi proteiinit ja käynnistäjäalueella sijaitsevat dna-jaksot, saattaa geenien ilmenemistä säädellä myös muualla kromosomissa sijaitsevat dna-jaksot. Nämä tehostaja-jaksot ovat tyypillisiä eukaryoottien transkription säätelyssä. Ne voivat sijaita kaukana, jopa tuhansien emäsparien päässä varsinaisesta transkription aloituskohdasta joko ylä- tai alavirtaan. Dna-molekyylin kiertyessä ja taipuessa lenkkimäiseksi ulottuu geenistä kaukanakin sijaitsevien säätelyjaksojen ja/tai niihin sitoutuneiden proteiinien vaikutus oikeaan kohteeseen. Tehostajat voivat sijaita kääntyneinä molemmin päin, joko 'alku-loppu'- tai 'loppu-alku'-suuntaisesti. Myös geenien ilmenemistä vaimentavia jaksoja on olemassa. Tehostajat ovat tärkeitä etenkin tietyssä kudoksessa tai kehitysvaiheessa ilmenevien geenien säätelyssä. Tehostajajaksojen sekä niihin sitoutuvien että muiden säätelevien proteiinien vuorovaikusten selvitys on nykyisen geenibiologisen tutkimuksen polttopisteessä. Proteiinien ja dna-jaksojen lisäksi geenien ilmenemisen säätely voi perustua m-rna- ja muihin lyhyisiin rn-molekyyleihin.Ensinnäkin transkription tuloksena syntyvän m-rna:n kestävyydestä johtuu, säilyykö tämä rna-molekyyli ollenkaan proteiineja valmistavan laitteiston käyttöön. Eukaryoottisoluissa on lisäksi kynnyksenä m-rna:n siirtyminen tumasta solulimaan, missä proteiineja valmistava laitteisto sijaitsee. Lisäksi lyhyet rna-molekyylit voivat toimia geenien ilmenemisen säätelijöinä. Rna-molekyyli voi hybridisoitua m-rna-molekyylin kanssa, mikäli ne ovat tarpeeksi komplementaarisia. Hybridi-rna hajoaa, eikä m-rna päädykään proteiinien valmistuksen ohjeeksi solulimaan. Rna-molekyylin sekundaarirakenteen muutos voi myös vaikuttaa molekyylin katkaisuun. Myös m-rna:n sekundaarirakenteen muutokset ovat keino geenien toiminnan säätelemiseksi. Rna-molekyylin sisäkkäiset emäspariutumiset aiheuttavat helposti lenkkirakenteita, joiden tiedetään vaikuttavan erityisesti transkription lopetukseen. Monilla bakteereilla ja joillakin eukaryooteillakin rna-molekyyliin muodostuva hiuspinniä muistuttava lenkkirakenne tarvitaan transkription lopetukseen - varsinainen lopetusjakson emäsjärjestys sinänsä ei ole niinkään olennainen. Prokaryooteille onkin ominaista, että translaatioon eteneminen riippuu transkription lopetusalueelle muodostuvista lenkkirakenteista.

51
Mutaatiot muuttavat perimän viestiä
Mutageenit Radioaktiivinen säteily, UV-säteily ja dna:n kanssa reagoivat kemikaalit Suurin osa aiheuttaa syöpää Mutaatiotyypit Pistemutaatio (yksittäinen emäs muuttuu) Kromosomimutaatio (kromosomin rakenne muuttuu) Häviämä eli deleetio Kahdentuma eli duplikaatio Säätelygeeneissä tapahtuvat mutaatiot voivat vaikuttaa useiden geenien toimintaan Kuva kirjan sivulta 72
 Kromosomimutaatio (kromosomin rakenne muuttuu) Häviämä eli deleetio. Kahdentuma eli duplikaatio. Säätelygeeneissä tapahtuvat mutaatiot voivat vaikuttaa useiden geenien toimintaan. Kuva kirjan sivulta 72.")
53
Sukusoluissa tapahtuva mutaatio siirtyy jälkeläisille ja luonnonvalinnan arvioitavaksi
Suurin osa tapahtuu somaattisissa soluissa eivät periydy Jos tapahtuu kehittyvissä ja runsaasti jakautuvissa kudoksissa, virhe periytyy solu ei pysty korjaamaan virhettä syöpä tai kehityshäiriö Jos mutaatio tapahtuu sukusolulinjan eli ituradan soluissa, se periytyy Mutaation vaikutukset: haitallinen neutraali hyödyllinen Kuva kirjan sivulta 72

54
Yhdistä kirjaimilla (a-h) merkitty käsite tarkimmin sitä vastaavaan numerolla (I-VIII) merkittyyn käsitteeseen. a) aaploidi b) diploidi c) polyploidi d) iturata e) replikaatiohaarukka f) silmukointi g) antikodoni h) perimä I) siirtäjä-rna II) dna:n kahdentuminen III) sukusolujen muodostama solulinja sukupolvesta toiseen IV) ihmisen somaattinen solu V) ihmisen sukusolu VI) 5n VII) yksilön kaikki dna VIII) esiaste-rna
 aaploidi b) diploidi c) polyploidi d) iturata e) replikaatiohaarukka f) silmukointi g) antikodoni h) perimä. I) siirtäjä-rna II) dna:n kahdentuminen III) sukusolujen muodostama solulinja sukupolvesta toiseen IV) ihmisen somaattinen solu V) ihmisen sukusolu VI) 5n VII) yksilön kaikki dna VIII) esiaste-rna.")
55
haploidi - V) ihmisen sukusolu
diploidi - IV) ihmisen somaattinen solu polyploidi - VI) 5n iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen replikaatiohaarukka - II) dna:n kahdentuminen f) silmukointi - VIII) esiaste-rna g) antikodoni - I) siirtäjä-rna h) perimä - VII) yksilön kaikki dna
 ihmisen somaattinen solu. polyploidi - VI) 5n. iturata - III) sukusolujen muodostama solulinja sukupolvesta toiseen. replikaatiohaarukka - II) dna:n kahdentuminen. f) silmukointi - VIII) esiaste-rna. g) antikodoni - I) siirtäjä-rna. h) perimä - VII) yksilön kaikki dna.")
56
4. Geenitekniikan perusmenetelmiä
1. Dna:n ja rna:n käsittely 2. Dna:n muokkaaminen 3. Dna:n ja rna:n tutkimusmenetelmiä 4. Geenien toiminnan tutkiminen 5. Tehtävät

57
Dna:n ja rna:n käsittely
Eristäminen Puhdistaminen Lähetti-rna:t voidaan vielä muuntaa niiden emäsjärjestystä vastaavaksi ns. komplementaariseksi dna:ksi Kuva kirjan sivulta 72

58
Dna:n muokkaaminen Katkaisuentsyymit Liittäjäentsyymit
Bakteerien luontainen puolustuskeino Pilkkovat dna:n halutusta kohdasta Samalla entsyymillä katkaistut dna:t voidaan liittää helposti yhteen Liittäjäentsyymit Liittää katkaistut juosteet yhteen Yhdistää myös eri alkuperää olevaa dna:ta Restriktioentsyymit ovat yhdistelmä-dna-tekniikan edellytys Eri restriktioentsyymit katkaisevat dna-molekyylin erilaisista emäsjaksoista. Sekä emäsjärjestys eli sekvenssi että sen pituus ovat tärkeitä. Restriktioentsyymit mahdollistavat nykyisen geenitekniikan sovelluksineen. Ne löydettiin alun perin bakteereista: bakteerit suojautuvat vierasta dna:ta vastaan hajottamalla sen. Restriktioentsyymejä käyttäen on mahdollista katkoa eri eilöstä peräisin olevaa dna:ta ja yhdistää katkotuista osista koostuva yhdistelmä-dna-molekyyli. Eniten dna:sta löytyy katkaisukohtia restriktioentsyymeille, jotka tunnistavat tiettyjä 4 emäksen jaksoja. On myös olemassa restriktioentsyymejä, jotka katkaisevat pidemmästä emäsjaksosta (5-, 6- tai 7-pilkkojat). On kuitenkin olemassa muuan entsyymi (terminaalideoksinukleotidyylitransferaasi), jonka avulla tylpät päät ovat muutettavissa tahmapäiksi. Tämän ligaasi-entsyymin avulla lisätään komplementaariset, yleensä A- ja T-hännät tylppiin päihin. Myös eräs faagiperäinen entsyymi, T4-faagin ligaasi, kykenee liittämään tylpät päät sellaisenaan. Niin sanottujen liittimien (engl. linker) avulla voi liittää myös tylpät päät toisiinsa tai johonkin muuhun molekyyliin. Liittimet ovat keinotekoisia lyhyehköjä dna-jaksoja, joihin voidaan sisällyttää restriktioentsyymien katkaisukohtia. Liittimiä käyttämällä voi samalla varmistaa, että siirrettävä geeni tulee asennetuksi oikein päin kohde-dna:han. Geenin ilmenemisen kannalta voi olla ratkaisevaa, että geenin alkupää tulee vastaanottajassa sijaitsevasta käynnistäjästä alavirtaan 5' - 3' -suuntaisesti. Sitä paitsi kaksi eri liitintä eivät voi takertua toisiinsa, joten näin vältytään lineaaristen juosteiden renkaiksi sulkeutumisilta.
. On kuitenkin olemassa muuan entsyymi (terminaalideoksinukleotidyylitransferaasi), jonka avulla tylpät päät ovat muutettavissa tahmapäiksi. Tämän ligaasi-entsyymin avulla lisätään komplementaariset, yleensä A- ja T-hännät tylppiin päihin. Myös eräs faagiperäinen entsyymi, T4-faagin ligaasi, kykenee liittämään tylpät päät sellaisenaan. Niin sanottujen liittimien (engl. linker) avulla voi liittää myös tylpät päät toisiinsa tai johonkin muuhun molekyyliin. Liittimet ovat keinotekoisia lyhyehköjä dna-jaksoja, joihin voidaan sisällyttää restriktioentsyymien katkaisukohtia. Liittimiä käyttämällä voi samalla varmistaa, että siirrettävä geeni tulee asennetuksi oikein päin kohde-dna:han. Geenin ilmenemisen kannalta voi olla ratkaisevaa, että geenin alkupää tulee vastaanottajassa sijaitsevasta käynnistäjästä alavirtaan suuntaisesti. Sitä paitsi kaksi eri liitintä eivät voi takertua toisiinsa, joten näin vältytään lineaaristen juosteiden renkaiksi sulkeutumisilta.")
61
Dna:n ja rna:n tutkimusmenetelmiä 1/2
Monistaminen PCR:n avulla tai liittämällä tutkittava alue bakteerin plasmidiin PCR Alukkeet Hyödynnetään mm. dna:n emäsjärjestyksen selvittämisessä eli sekvensoinnissa ja yksilöiden tunnistuksessa Elektroforeesi Käytetään erimittaisten dna- ja rna-palasten havaitsemiseen Geenin dna:n erojen tutkiminen Dna:n hybridisaatio Dna:n juosteiden irrottaminen toisistaan kuumentamalla PCR-menetelmän (engl. polymerase chain reaction) avulla tiettyjä, haluttuja dna-jaksoja voidaan monistaa, kun käytetään sopivia alukkeita. Esimerkiksi geenitutkimuksessa voidaan verinäytteistä eristetystä dna:sta ’rikastaa’ tiettyä geenilokusta. Näin voidaan tutkia vaikkapa eri alleelien osuuksia väestössä. PCR:llä voidaan myös ’rikastaa’ dna:ta esimerkiksi rikospaikalle jääneestä veritahrasta tai muusta eritetahrasta. Jopa muumioiden dna:ta on PCR:llä monistettu tutkimuksia varten. Elektroforeesissa dna-, rna, tai proteiinimolekyylejä tai niiden osia voi erottaa koon mukaan. Elektroforeesilaitteistossa on kaksi elektrodia, katodi ja anodi, joita yhdistää sähköä kantavana väliaineena elektrolyyttiliuos. Tutkittavat molekyylit liikkuvat sähkökentässä virran mukana. Etenkin nukleiinihappomolekyylien kantajana käytetään geelejä. Elektroforeesiajon aikana kooltaan pienimmät molekyylit ehtivät liikkua geelissä kaikkein kauimmas.
 avulla tiettyjä, haluttuja dna-jaksoja voidaan monistaa, kun käytetään sopivia alukkeita. Esimerkiksi geenitutkimuksessa voidaan verinäytteistä eristetystä dna:sta ’rikastaa’ tiettyä geenilokusta. Näin voidaan tutkia vaikkapa eri alleelien osuuksia väestössä. PCR:llä voidaan myös ’rikastaa’ dna:ta esimerkiksi rikospaikalle jääneestä veritahrasta tai muusta eritetahrasta. Jopa muumioiden dna:ta on PCR:llä monistettu tutkimuksia varten. Elektroforeesissa dna-, rna, tai proteiinimolekyylejä tai niiden osia voi erottaa koon mukaan. Elektroforeesilaitteistossa on kaksi elektrodia, katodi ja anodi, joita yhdistää sähköä kantavana väliaineena elektrolyyttiliuos. Tutkittavat molekyylit liikkuvat sähkökentässä virran mukana. Etenkin nukleiinihappomolekyylien kantajana käytetään geelejä. Elektroforeesiajon aikana kooltaan pienimmät molekyylit ehtivät liikkua geelissä kaikkein kauimmas.")
62
Dna:n ja rna:n tutkimusmenetelmiä 2/2
Koetin: Merkitään leimaamalla fluoresoivalla väriaineella tai radioaktiivisella merkillä Koetin ja tutkittava kohde pariutuvat keskenään Sekvensointi Dna:n emäsjärjestyksen selvittäminen Sekvenssit tallennetaan tietokantoihin 4. Sekvenointimenetelmillä saadaan tietoa emäsjärjestyksestä Sekvenointi tarkoittaa dna:n emäsjärjestyksen selvittämistä. Automaattiset sekvenointilaitteet mahdollistivat ihmisgenomin kartoituksessa työn etenemisen, joka muuten olisi ollut todella hidasta. Sekvenointi-menetelmiä on kaksi: Sangerin menetelmä ja Maxam-Gilbertin menetelmä

64
Geenien toiminnan tutkiminen
Geeni toimii, kun se tuottaa lähetti-rna:ta Geeniekspressiota tutkitaan mittaamalla lähetti-rna-molekyylien määrä Dna-siru lasista tai muovista valmistettu pieni levy, johon on kiinnitetty DNA-palasia Voidaan tutkia ympäristötekijöiden, ravintoaineiden tai lääkehoitojen vaikutusta geenien toimintaan Voidaan havaita yksilöiden välisiä eroja dna:n emäsjärjestyksessä Mikrosirutekniikalla tutkitaan geenien ilmenemistä Mikrosirutekniikka mahdollistaa sen, että yhtäaikaa voidaan tutkia jopa geenien ilmenemistä eri kudoksista tai solukoista otetuissa näytteissä. Sirun mikroskooppilasilla on tuhansia geenikoettimia. Myynnissä olevissa siruissa on erilaisia yhdistelmiä näistä koettimista, siru voi olla esimerkiksi kehitetty syöpägeenien tutkimista varten. Näihin koettimiin hybridisoidaan tutkittavista näytteistä eristettyä c-dna:ta, joka on leimattu fluorisoivilla väriaineilla. Nykyisin käytetään paljon oligonukleotideista (25-70 emästä) valmistettuja siruja. Etuna c-dna-klooneihin verrattuna on se, että näin voidaan tunnistaa niin yksittäisiä geenejä kuin geenin muunnoksiakin. Näin voidaan tutkia esimerkiksi vaihtoehtoisen silmikoinnin tai polyadenylaation merkitystä. Siruanalysissä siru luetaan laser-pohjaisella mikroskoopilla.
 valmistettuja siruja. Etuna c-dna-klooneihin verrattuna on se, että näin voidaan tunnistaa niin yksittäisiä geenejä kuin geenin muunnoksiakin. Näin voidaan tutkia esimerkiksi vaihtoehtoisen silmikoinnin tai polyadenylaation merkitystä. Siruanalysissä siru luetaan laser-pohjaisella mikroskoopilla.")
66
Nimeä molekyyli, menetelmä tai laite.
dna:n emäsjärjestyksen määrittäminen dna:n monistaminen erimittaisten dna-jaksojen erottaminen toisistaan geenien toiminnan tutkiminen eri eliöiden dna:ta sisältävä molekyyli

Samankaltaiset esitykset
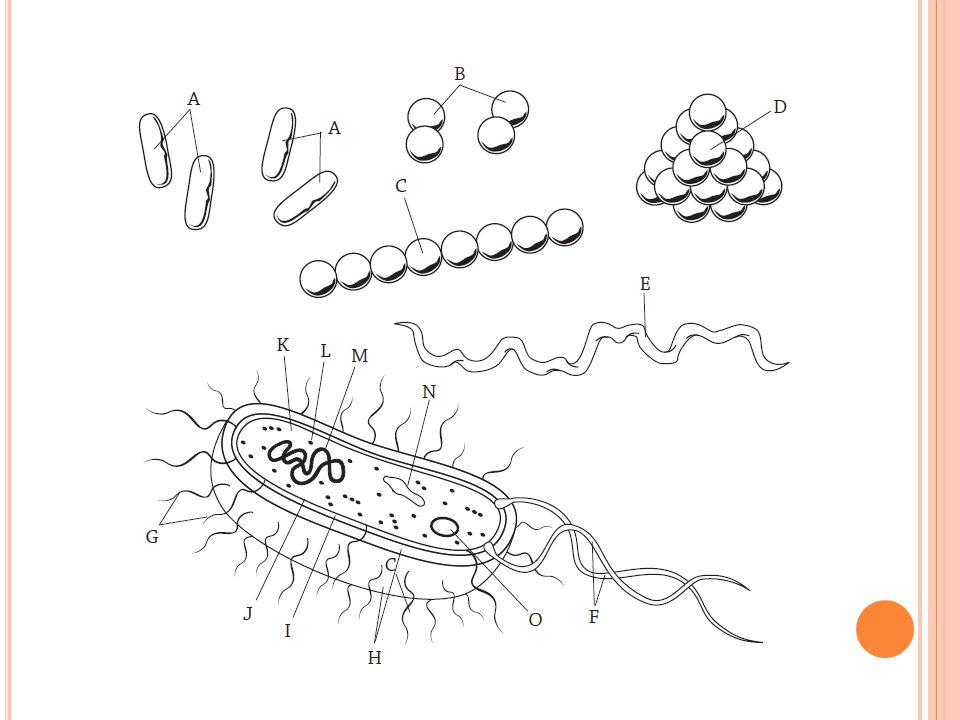
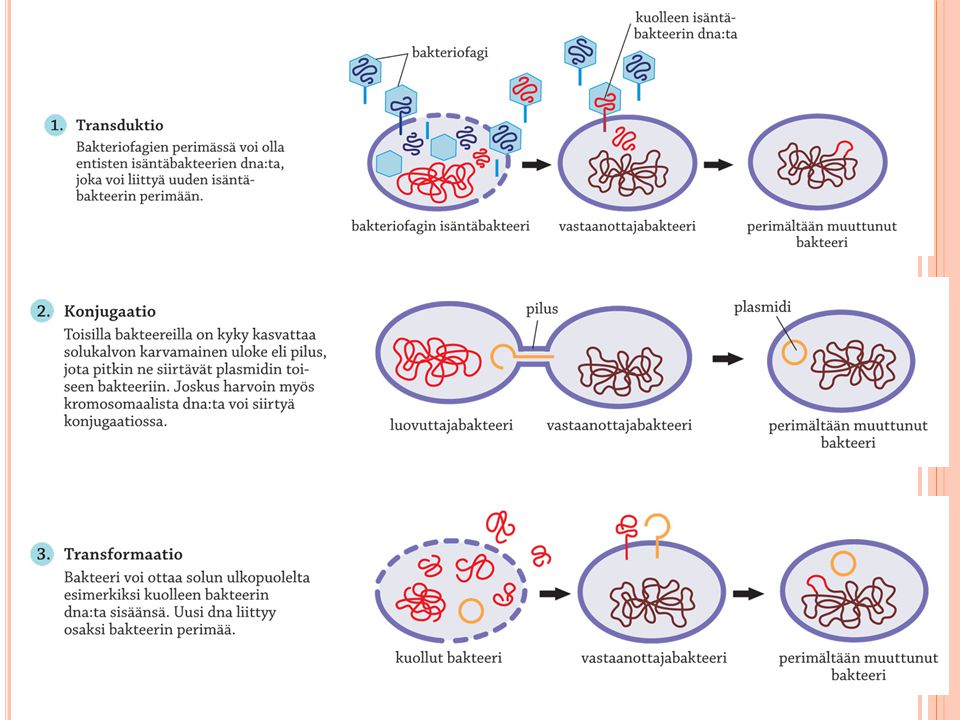
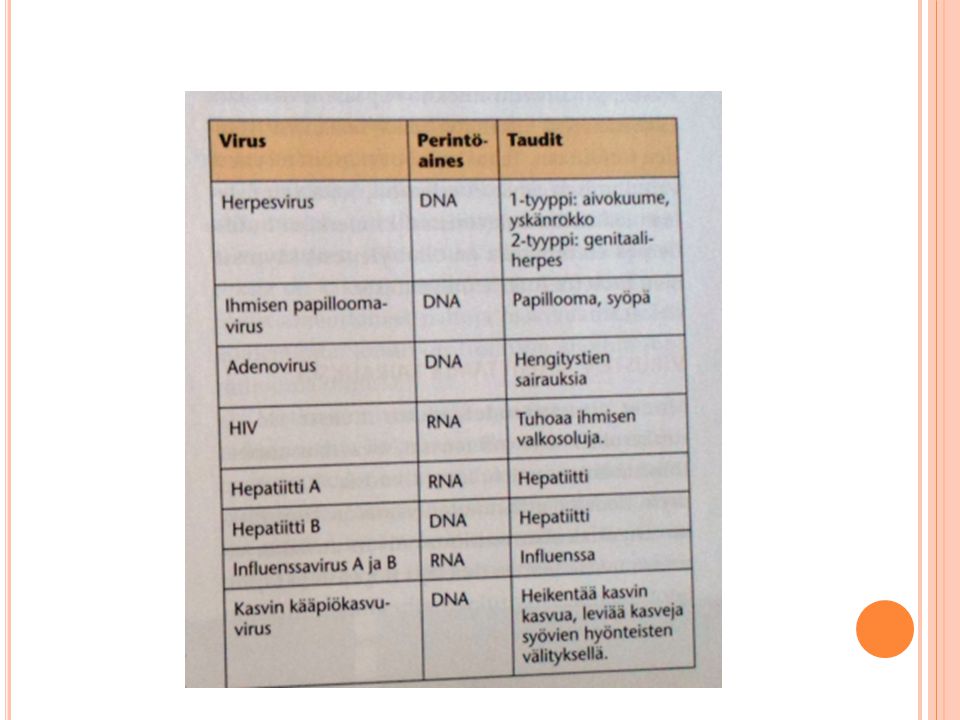
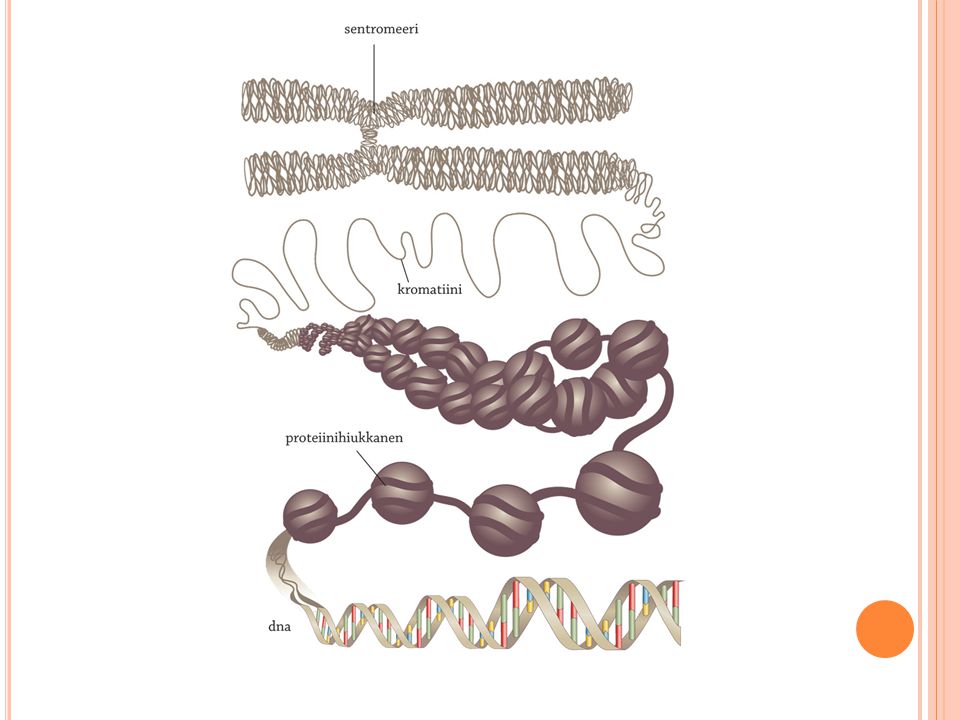

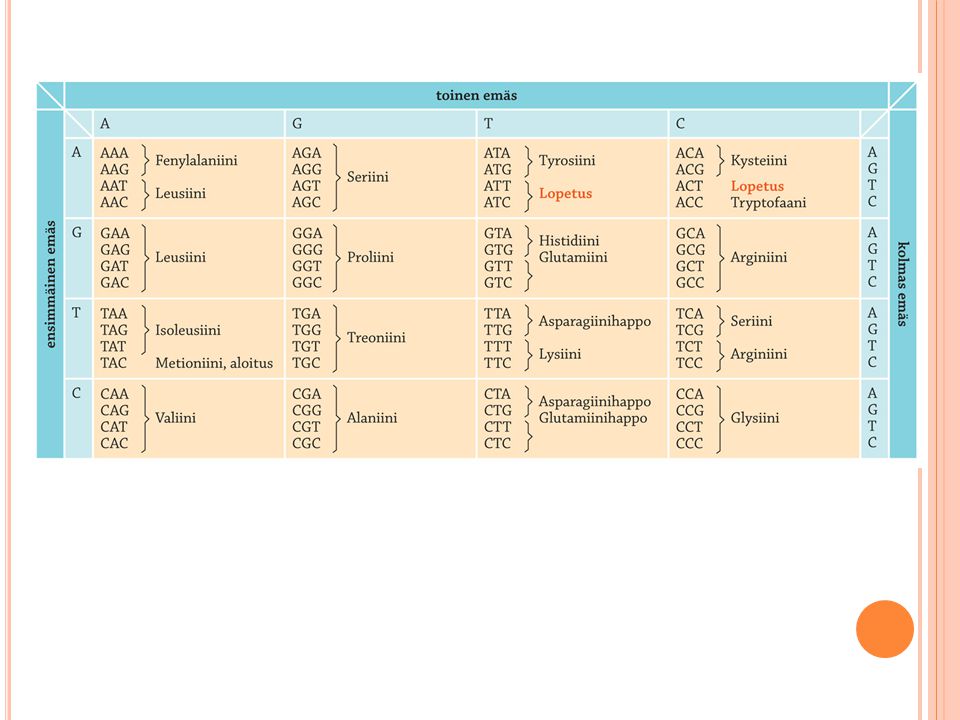
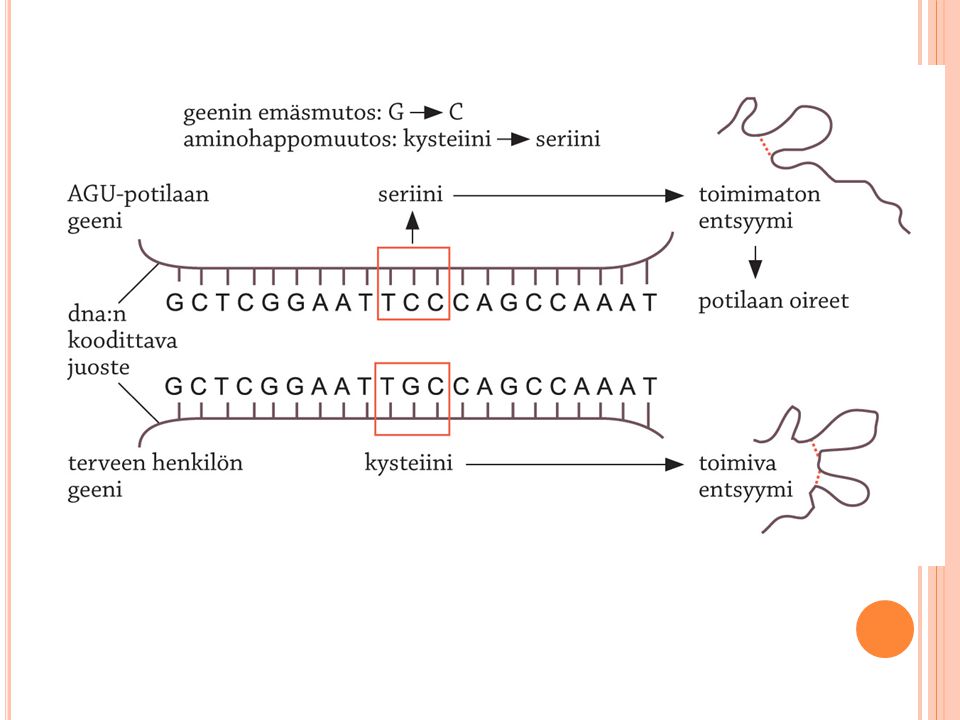
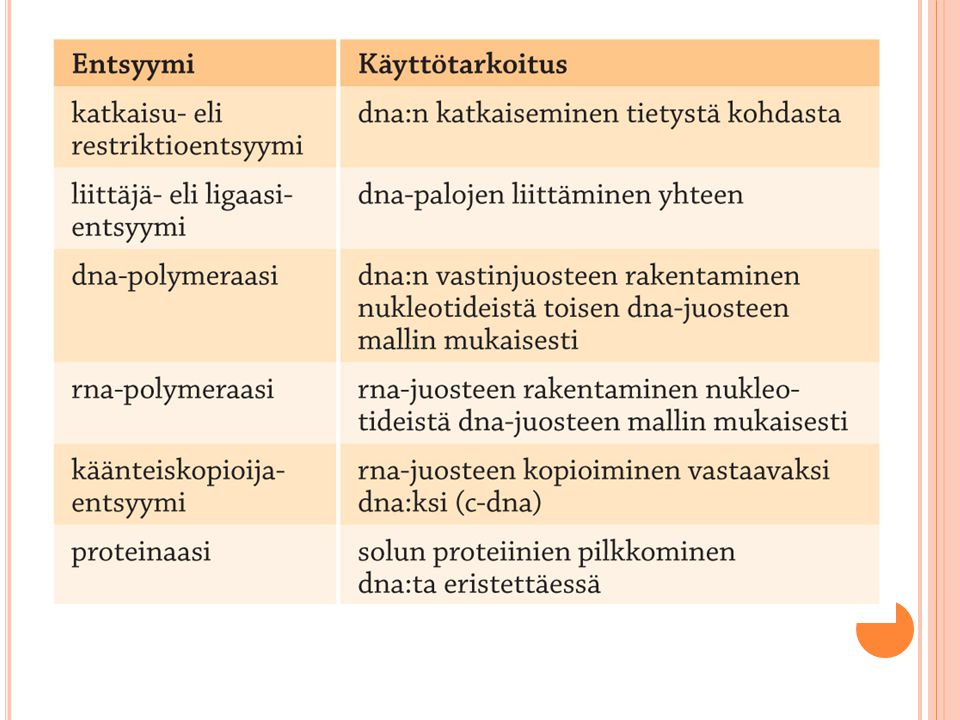
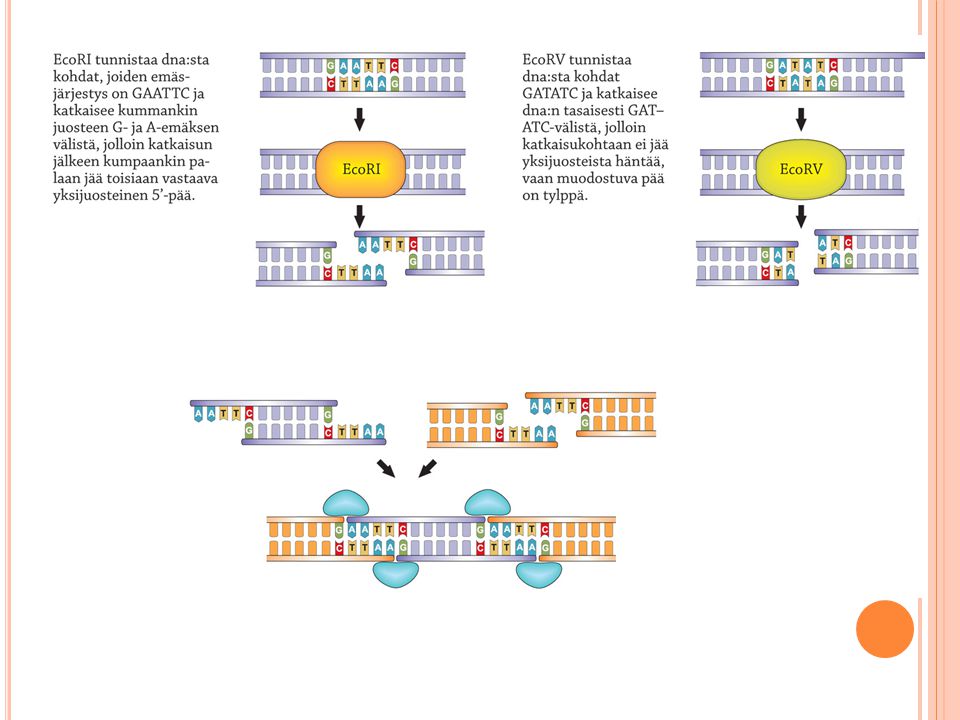
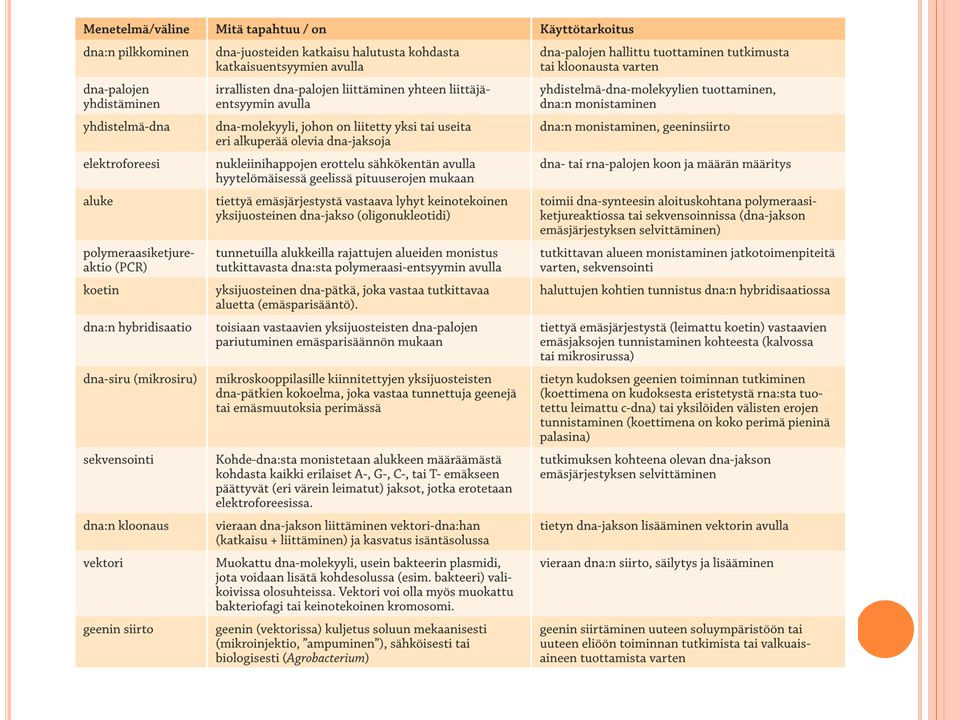
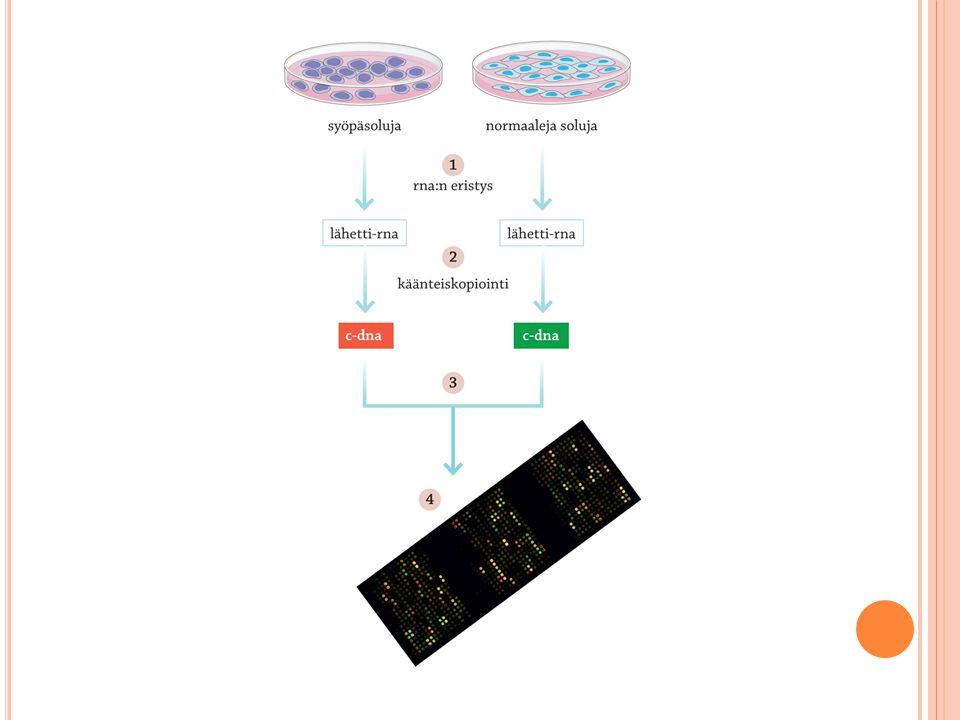








.>")










